XiaoMi-AI文件搜索系统
World File Search SystemPRMT5

在SARS-COV-2
非霍奇金淋巴瘤(NHL)是在淋巴组织中产生的一组血液癌,通常会影响人类和狗。蛋白精氨酸甲基转移酶5(PRMT5)是一种催化精氨酸残基的对称二甲基化的酶,在人类固体和血液系统恶性肿瘤中均过表达且失调。在人淋巴瘤中,PRMT5是已知的恶性转化和肿瘤发生的驱动因素,但是尚未探索PRMT5在犬淋巴瘤中的表达和作用。探索犬淋巴瘤是与人淋巴瘤的有用比较,同时将PRMT5作为两者中的有理治疗靶标的,我们表征了犬淋巴瘤组织,原发性淋巴样生物的PRMT5的表达模式,以及犬淋巴瘤衍生的细胞系。PRMT5的抑制导致了抑制和诱导凋亡,同时选择性降低了对称二甲基精氨酸(SDMA)(SDMA)和组蛋白H4精氨酸3对称二甲基化的全局标记。,我们通过途径富集分析进行了ATAC测序和基因表达微阵列,以表征全基因组可及性的全基因组变化和PRMT5抑制后犬淋巴瘤细胞系的全转录组变化。这项工作将PRMT5验证为犬淋巴瘤的有前途的治疗靶标,并支持继续使用自发发生的犬淋巴瘤模型,用于临床前PRMT5抑制剂治疗人类NHL。

全基因组 R 环分析确定了 DDX5、XRN2 和 PRMT5 在 DNA/RNA 杂交分辨率中的独特作用
研究表明,DDX5、XRN2 和 PRMT5 可以在少数基因组位点上解析 RNA 聚合酶 II 转录终止位点处的 DNA/RNA 杂合体 (R 环)。在此,我们使用经典的 DNA/RNA 免疫沉淀和高通量测序 (DRIP-seq) 对受 DDX5、XRN2 和 PRMT5 调控的位点进行全基因组 R 环定位。我们在缺乏 DDX5、XRN2 和 PRMT5 的 U2OS 细胞中观察到转录位点处数百到数千个 R 环增益和丢失。R 环增益是位于基因富集区域的高度转录基因的特征,而 R 环丢失则在低密度基因区域观察到。DDX5、XRN2 和 PRMT5 在转录终止位点共享许多 R 环增益位点,这与它们在 RNA 聚合酶 II 转录终止中的协调作用一致。 DDX5 缺失的细胞在转录起始位点附近具有独特的 R 环增益峰,这些峰与 siXRN2 和 siPRMT5 细胞的 R 环增益峰不重叠,表明 DDX5 在转录起始中发挥独立于 XRN2 和 PRMT5 的作用。此外,我们观察到 siDDX5、siXRN2 和 siPRMT5 细胞中基因转录起始位点附近某些位置的累积 R 环导致反义基因间转录。我们的研究结果确定了 DDX5、XRN2 和 PRMT5 在 DNA/RNA 杂交调控中的独特和共同作用。

免疫治疗
a)(左)PRMT5纳米底测定的示意图以及MTA或SAM对示踪剂结合的影响,改编自参考文献2。(右)HCT116等生成对中的PRMT5纳米杆。细胞用指定剂量的IDE397预处理23小时,并测量对示踪剂结合的影响(左)。预先处理IDE397(23小时),然后添加MRTX1719持续2小时(右)。b)HCT116 wt(顶行)或mtap-/ - (底行)中IDE397的全剂量矩阵和PRMT5抑制剂;热图中显示的明显目标占用率。由10µM GSK3326595(探针母体分子) + 100nm的IDE397预处理前的MBRET比定义了100%的明显占用(最大探针位移)。0%的明显占用率仅代表DMSO。因此,100%明显的目标占用率代表PRMT5抑制剂与PRMT5的最大结合。
Adam Yaari
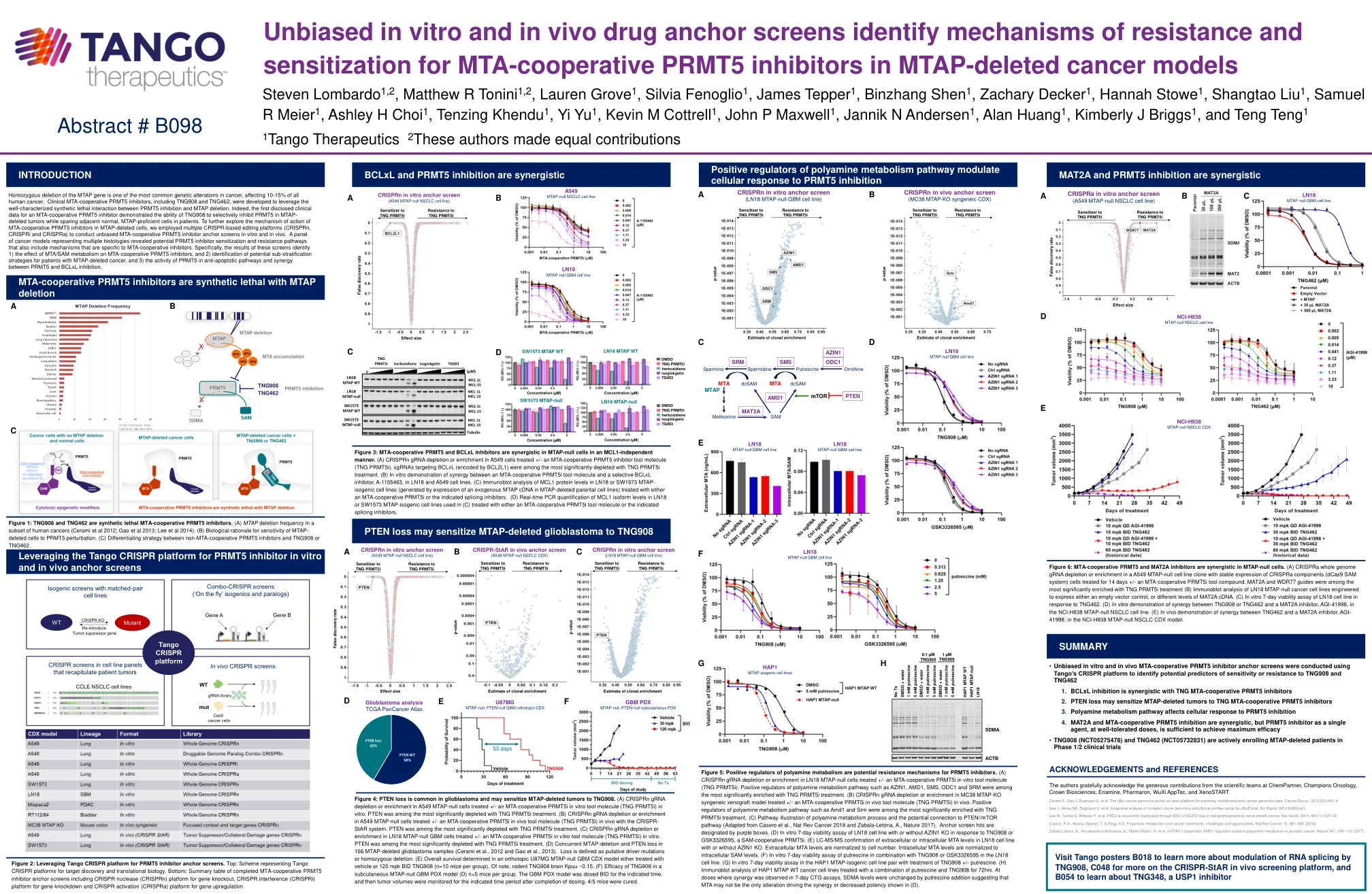
MTAP基因的纯合缺失是癌症中最常见的遗传改变之一,影响了所有人类癌症的10-15%。开发了包括TNG908和TNG462在内的临床MTA合并PRMT5抑制剂,以利用PRMT5抑制和MTAP缺失之间充分表征的合成致死相互作用。的确,第一个披露的MTA合作性PRMT5抑制剂的临床数据证明了TNG908在MTAP删除的肿瘤中有选择性地抑制PRMT5的能力,同时在患者中保留邻近的正常,MTAP-Profofoffic-Profofoffic-MTAP-Profofoffoffient。为了进一步探索MTA骨骼删除细胞中MTA合件性PRMT5抑制剂的作用机理,我们采用了多个基于CRISPR的编辑平台(CRISPRN,CRISPRI和CRISPRA)来进行无偏MTA合件性PRMT5抑制剂锚固筛网,并在In In In In In Intro和Vivo中进行。代表多种组织学的癌症模型揭示了潜在的PRMT5抑制剂敏化和耐药途径,这些途径还包括特定于MTA合并抑制剂的机制。Specifically, the results of these screens identify 1) the effect of MTA/SAM metabolism on MTA-cooperative PRMT5 inhibitors, and 2) identification of potential sub-stratification strategies for patients with MTAP-deleted cancer, and 3) the activity of PRMT5 in anti-apoptotic pathways and synergy between PRMT5 and BCLxL inhibition.

通过MTAP - / - 肿瘤中MAT2A和PRMT5组合抑制MAT2A和PRMT5的深层和耐用抗肿瘤活性的机械基础
本演讲中的某些陈述和随附的口头评论是前瞻性陈述。这些陈述涉及未来事件或Ideaya Biosciences,Inc。(“公司”)的未来财务绩效,并涉及已知和未知的风险,不确定性和其他因素,这些风险可能会导致公司或其行业的实际结果,活动水平,绩效或其行业的水平与任何前瞻性陈述表达或暗示。在某些情况下,可以通过术语来识别前瞻性陈述,例如“可能”,“意志”,“可能”,“将”,“应该”,“期望”,“预期”,“预期”,“预期”,“相信”,“估计”,“预测”,“预测”,“潜在”,“潜在”或其他可比较的术语。公司将这些前瞻性陈述基于其当前的期望,假设,估计和预测。虽然公司相信这些期望,假设,估计和预测是合理的,但前瞻性陈述仅是预测,涉及已知和未知的风险和不确定性,其中许多超出了公司的控制。这些和其他重要因素可能导致实际结果,绩效或成就与这些前瞻性陈述所表达或暗示的因素有重大不同。本演示文稿中的前瞻性语句仅在此日期开始。除法律要求外,公司没有承担任何义务,也不打算更新这些前瞻性陈述,或者将这些陈述符合实际结果或公司期望的变化。除了历史事实陈述以外的所有陈述都可以视为前瞻性,包括对临床活动概况,潜在的临床益处和公司临床计划的潜在优势的期望;初步临床试验结果转化为将来的临床试验结果;临床试验的入学;在转移性卵巢黑色素瘤中对Darovasertib和Crizotinib组合评估的2/3期临床试验是否将被美国食品药品监督管理局(“ FDA”)视为注册试验;该公司计划的潜在可寻址患者群体;关于公司目标发现平台或新目标验证工作的任何期望,以创造研发计划的机会;财务信息,市场机会,现金跑道或盈利能力的任何预测,包括估计的运营资金到2028年;关于历史结果的任何陈述,这些陈述可能暗示了公司业务的趋势;针对开发计划或未来运营的管理计划,策略和目标的任何陈述;关于临床前研究,临床开发,监管文件,制造或释放数据时机的任何陈述;关于未来事件,潜在市场或市场规模,技术发展或收到现金里程碑,期权行使费用或特许权使用费的任何期望或信念的陈述;以及提到任何项目的基础假设的任何陈述。此类风险和不确定性包括药物开发过程中固有的不确定性,包括公司的计划的早期阶段,设计和进行临床前和临床试验的过程,严重的不良事件,不良不良事件,不良的副作用,不良的副作用,不良的副作用或药物开发的意外特征知识产权以及其他可能影响现金足够资金运营的事项。以进一步描述可能导致实际结果与这些前瞻性陈述中表达的结果不同的风险和不确定性,以及与公司业务有关的风险以及与美国证券交易委员会(SEC和SEC)的定期申请(包括在10月31日的10年代终止的年度日期为10年代)的年度报告(包括其年度报告)的年度报告,并在20223年6月3日报告中,请参阅公司的定期申请,包括该表格的年度报告。向SEC提交的当前或周期性报告。

Prism合作者幻灯片的资源页面
●针对MTA合并性PRMT5抑制剂(在第1/2阶段试验中)的基因表达,蛋白质组学,shRNA敲低和广泛的重新利用数据集相关性,据报道优先针对MTAP-MTAP-浸没细胞活性。●分别具有PRMT5和MTAP的化合物靶标和预期的生物标志物是最强的命中率之一,与Prism中的复合响应相关

繁殖 - Bioscientifica
范可尼贫血 (FA) DNA 损伤反应 (DDR) 通路调节重要的细胞过程,例如 DNA 复制、细胞周期控制和 DNA 损伤修复。本文我们表明,FANCD2 是 FA DDR 通路的关键成员,它与生殖细胞特异性 Prmt5/piRNA 通路的几个重要成分相互作用,这些通路协调对转座因子 (TE) 的抑制。通过使用标记纯原始生殖细胞 (PGC) 群体的 Pou5f1 -eGFP 报告小鼠,我们证明 FA 缺乏会导致 TE 的抑制解除、PGC 耗竭以及精子发生和卵子发生缺陷。Fancd2-KO PGC 表现出过度的 DNA 损伤并加剧细胞凋亡。从机制上看,我们观察到在 Fancd2-KO ; Pou5f1 -eGFP 和 Fanca-KO ; Pou5f1 -eGFP 胚胎的 E10.5 PGC 中,PRMT5 催化的 H2A/H4R3me2s 标记在 LINE1 TE 上显著减少。此外,我们利用 Fancd2-KI 模型表明,在 WT PGC 中,FANCD2 和 PRMT5 共同占据了 LINE1 的启动子,而在 FA 缺陷型(Fanca-KO)PGC 中,这种共同占据消失了。这些结果表明,FA 通路参与了早期 PGC 中的 TE 抑制,可能通过一种涉及 FANCD2 促进的、PRMT5 催化的抑制性 H2A/H4R3me2s 标记的机制来实现。生殖 (2020) 159 659–668

靶向MYC与表观遗传调节剂结合使用,在MLLR白血病中诱导协同的抗白血病作用,并同时提高免疫力
mll重排(MLL R)白血病与预后不良和对常规疗法的反应有限有关。此外,化学疗法会导致严重的侧面影响,并严重受到免疫系统的损害。因此,必须识别新型治疗策略。最近,我们通过使用簇状的定期插入的短篇小学重复序列(CRISPR)/cas9在CD34 +细胞中诱导CD34 +细胞中的染色体重排,开发了人类MLL RR白血病模型。该MLL R模型的真实性模仿患者白血病细胞,可用作新型治疗策略的平台。我们模型的 RNA测序揭示了MYC是促进造成发生的最重要的关键驱动因素之一。 然而,在临床试验中,BRD4抑制剂JQ-1导致间接阻断MYC途径仅显示适度的活动。 我们和其他人以前报道说,靶向MAT2A或PRMT5的表观遗传药物促进了MLL R细胞中的细胞死亡。 因此,我们将这些药物与JQ-1结合使用,从而增强了抗白血病效应。 更重要的是,我们发现T,NK和INKT细胞的激活,免疫调节细胞因子的释放以及抑制剂治疗后PD-1/PD-L1轴的下调导致细胞毒性提高。 总而言之,MYC和MAT2A或PRMT5的抑制作用驱动了MLL RL白血病的强大协同抗白血病活性。 此外,在组合抑制剂治疗后同时激活免疫系统,从而进一步提高了治疗效率。RNA测序揭示了MYC是促进造成发生的最重要的关键驱动因素之一。然而,在临床试验中,BRD4抑制剂JQ-1导致间接阻断MYC途径仅显示适度的活动。我们和其他人以前报道说,靶向MAT2A或PRMT5的表观遗传药物促进了MLL R细胞中的细胞死亡。因此,我们将这些药物与JQ-1结合使用,从而增强了抗白血病效应。更重要的是,我们发现T,NK和INKT细胞的激活,免疫调节细胞因子的释放以及抑制剂治疗后PD-1/PD-L1轴的下调导致细胞毒性提高。总而言之,MYC和MAT2A或PRMT5的抑制作用驱动了MLL RL白血病的强大协同抗白血病活性。此外,在组合抑制剂治疗后同时激活免疫系统,从而进一步提高了治疗效率。

抗蛋白精氨酸甲基转移酶5(PRMT5)抗体的鉴定和验证是全身性硬化的新生物标志物(SSC)
抽象目标急性前葡萄膜炎(“葡萄膜炎”)是一种常见的轴向脊椎关节炎(AXSPA)骨骼外骨骼外表现。白介素(IL)-17与其发病机理有关,但是,在葡萄膜炎管理中抑制IL-17A的证据相互矛盾。我们报告了接受Bimekizumab(BKZ)的患者的葡萄膜炎发病率的合并分析,这是一种单克隆IgG1抗体,从2B/3阶段试验中选择性地抑制IL-17F,除IL-17A外,还可以抑制IL-17F。方法数据,为移动1(NCT03928704;非放射线学AXSPA),并进行移动2(NCT03928743;放射线学AXSPA)试验。数据分别汇总了在BE移动试验中至少使用一个BKZ剂量的患者及其持续的开放标签扩展(OLE; NCT04436640)的数据,以及2B期Be Agile试验(NCT02963506;放射线摄影型AXSPA)及其正在进行的OLE(NCT033555555573)。葡萄膜炎率和暴露调整后的发病率(EAIR)/100患者年(PYS)。Results In the BE MOBILE 1 and 2 double-blind treatment period, 0.6% (2/349) of patients receiving BKZ experienced uveitis vs 4.6% (11/237) receiving placebo (nominal p=0.001; EAIR (95% CI): 1.8/100 PYs (0.2 to 6.7) vs 15.4/100 PYs (95% CI 7.7 to 27.5)).在葡萄膜炎史的患者中,接受BKZ的患者较低(6.2/100 PYS(95%CI 0.2至34.8); 1.9%)vs安慰剂(70.4/100 PYS(95%CI 32.2至133.7); 20.0%; 20.0%;名义P = 0.004)。在2b/3阶段池中(n = 848; BKZ暴露:2034.4 PYS),EAIR保持较低(1.2/100 PYS(95%CI 0.8至1.8))。结论Bimekizumab是一种双IL-17A/F抑制剂,可能对AXSPA患者赋予葡萄膜炎的保护作用。

蛋白精氨酸甲基转移酶5(PRMT5)作为非霍奇金淋巴瘤自发犬模型中的候选治疗靶标
凝结材料和冷凝物质物理中心(IFIMAC),马德里大学,马德里大学28049大学,西班牙B物理学和天文学系,奥尔胡斯大学,阿尔胡斯大学,阿尔胡斯C 80 0,丹麦·塞斯纳(Dev) 28049,西班牙D物理研究所,里约热内卢大学,邮政信箱68528,里约热内卢,RJ 21941-972,巴西兼物理系,埃斯皮里托·桑托大学联邦大学,维多利亚大学,维多利亚大学,ES 29075-910,29075-910无机化学,化学科学高级研究所(IADCHEM)和凝分物理学中心(IFIMAC),马德里28049年,马德里28049,HALBA Synchrotron,La llum Carrer 2-26凝结材料和冷凝物质物理中心(IFIMAC),马德里大学,马德里大学28049大学,西班牙B物理学和天文学系,奥尔胡斯大学,阿尔胡斯大学,阿尔胡斯C 80 0,丹麦·塞斯纳(Dev) 28049,西班牙D物理研究所,里约热内卢大学,邮政信箱68528,里约热内卢,RJ 21941-972,巴西兼物理系,埃斯皮里托·桑托大学联邦大学,维多利亚大学,维多利亚大学,ES 29075-910,29075-910无机化学,化学科学高级研究所(IADCHEM)和凝分物理学中心(IFIMAC),马德里28049年,马德里28049,HALBA Synchrotron,La llum Carrer 2-26