XiaoMi-AI文件搜索系统
World File Search SystemRNA

脂质纳米颗粒用于RNA递送
在非病毒载体中,脂质纳米摩析被认为是递送RNA thera peutics的金标准。脂质纳米颗粒在RNA递送中的成功,并批准了三种用于人使用的产品,已激发了对不同病理学的RNA疗法的进一步研究。这需要解码病理的细胞内过程并将输送系统定制到目标组织和细胞。脂质纳米分离器形态的复杂性源于脂质成分的组装,可以通过各种能够驱动纳米颗粒与所需组织的纳米颗粒形成的方法引起的脂质成分。在其他情况下,可以将预成型的纳米颗粒与RNA混合,以诱导自组装和结构重新构造成RNA负载的纳米颗粒。在这篇综述中,最相关的脂质纳米传动器及其RNA传递的潜力是根据组装机理和粒子结构描述的。

RNA的开发和特定疾病调节... 血浆中成纤维细胞生长因子与颈动脉斑块新生血管形成:一项初步研究 低肌肉质量指数与拉丁美洲人群中的2型糖尿病风险有关:横断面研究 西班牙裔,黑人和白人的邻里障碍和2型糖尿病差异 神经炎症中的先天淋巴样细胞 对代偿性心力衰竭的门诊管理的护理系统 dual-... 上定量参数的诊断值 足球杂耍学习对执行功能和大脑功能连接的影响 补充藏红花对糖尿病血糖结果的影响:系统评价和荟萃分析 I型干扰素信号传导,认知和神经退行性19:在机械致病模型上更新,对阿尔茨海默氏病的影响
替代剪接是一个复杂的基因调节过程,它通过重新安排未成熟前MRNA转录本的内含子和外显子和外显子来区分自身。这个过程在增强基因组的转录组和蛋白质组学多样性中起着至关重要的作用。替代剪接已成为一种关键机制,该机制是在心脏发育和心血管疾病发展过程中的复杂生物学过程的关键机制。在相关生理过程中,在重要基因的调节中以协同或拮抗的方式涉及多个替代剪接因子。值得注意的是,圆形RNA直到最近才引起了其特异性表达模式和调节功能的关注。这种兴趣的复兴促使对该主题进行了重新评估。在这里,我们概述了我们当前对替代剪接机制的理解以及替代剪接因子在心血管发育中的替代剪接因子的调节作用,以及不同心血管疾病的病理学过程,包括心肌病,心肌梗死,心力衰竭,心脏失败,心脏失败,心脏失败和动脉粥样硬化。

G2 DNA/RNA增强子
G2 DNA/RNA增强子可以方便地使用,尤其是尤其是粘土中需要最佳的DNA和/或RNA提取产率时。G2 DNA/RNA增强子的主要功能是减轻抑制性DNA-粘土颗粒的形成。G2 DNA/RNA增强子增加了粘土的微生物DNA和RNA产量 - 至少2-10倍。G2 DNA/RNA增强剂应与标准化提取方法或用于从土壤和粘土中提取DNA和RNA的商业试剂盒结合使用。建议在-20至25°C处进行存储和稳定性存储。保持干燥。质量控制G2 DNA/RNA增强子进行污染活性,没有核酸内核酸酶活性,缺口活性,外切核酸酶活性或RNase活性的痕迹。此外,在难以提取的矩阵中,对G2 DNA/RNA增强子进行了功能测试。套件组件Ampliqon G2 DNA/RNA增强子冻结干燥的G2 DNA/RNA增强剂和2 mL管中的1.4 mM珠。协议使用G2 DNA/RNA增强子时,该方案是DNA和RNA提取的指南。G2 DNA/RNA增强子必须使用提取套件施加。程序:将0.25克土壤样品添加到G2 DNA/RNA增强器管中。应用您的DNA或RNA隔离套件。例如Dneasy Powersoil Pro Kit。o如果套件的珠珠管中包含裂解缓冲液,请将此裂解缓冲液转移到G2管上,并丢弃现在空的套件的珠珠管。

Quick-DNA/RNA FFPE套件
产品描述Quick-DNA/RNA™FFPE试剂盒为一个简单可靠的方法,用于从一个单独的福尔马林膜状帕片蛋白嵌入(FFPE)组织样品中的两个单独的部分中的一种洗脱酸或DNA和RNA中的总核酸分离(DNA/RNA)。该产品的独特化学成分已被优化,以最大程度地恢复DNA和大小和小RNA物种。只需使用脱氧蛋白溶液将组织脱氧化,使用蛋白酶K消化,进行热化学交联,然后使用Zymo-Spin™柱技术进行纯化。结果是不含DNA的高质量DNA和总RNA(包括小RNA),可以用于RT/QPCR,杂交,测序等。
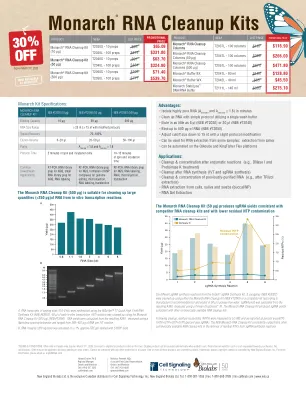
Monarch®RNA清理套件
使用Monarch RNA清洁套件(NEB#T2040)或竞争对手试剂盒(根据制造商的建议)清理了Engen®SgrNA合成试剂盒(NEB#E3322)的六种不同的SGRNA合成反应(NEB#E3322)(NEB#E3322),并在50μL的50μLNutece-Free Free Water中进行了体重。SGRNA产量是根据由Trinean Dropsense™16进行测量的结果A260计算的。君主RNA清理套件产生的SGRNA产生与其他市售RNA清理套件一致的。清理后,通过LC-MS测量残留核苷酸(NTP),并报告为面积为NTPS百分比(RATP+RCTP+RGTP+RUTP)/SGRNA百分比。NEB Monarch RNA清理套件始终优于其他市售的RNA清洁套件,从而从SGRNA合成反应中去除残留的NTP。

使用合成 RNA 作为 RNA 直接测序中 poly(A) 长度推断的基准
多聚腺苷酸化是一个动态过程,在细胞生理学中非常重要。Oxford Nanopore Technologies 的直接 RNA 测序提供了一种对全长 RNA 分子进行测序以及对转录组和表观转录组进行分析的策略。目前有几种可用于 poly(A) 尾长估计的工具,包括 tailfindr 和 nanopolish 等成熟工具,以及两种较新的深度学习模型:Dorado 和 BoostNano。然而,这些工具的准确性与金标准数据集的基准测试有限。在本文中,我们使用合成 RNA 标准(Sequins)评估了四种 poly(A) 估计工具,这些标准具有已知的 poly(A) 尾长,并提供了一种衡量 poly(A) 尾长估计准确性的有效方法。所有四种工具生成的平均尾长估计值都在正确值的 12% 以内。总体而言,由于 Dorado 运行时间相对较快、变异系数低并且易于使用且可与碱基调用集成,因此被推荐作为首选方法。

5'-CAP RNA/SAM模拟偶联物作为病毒RNA CAP 2'-O-O-甲基转移酶的双底物抑制剂
试剂和条件:; (a)二苯基磷酸盐 /吡啶,40°C,35分钟; (b)0.1m茶具,pH 8,40°C,45分钟; (c)咪唑,CBRCL 3,BSA,ET 3 N,MECN,40°C,1 h; (d)对于gppporn:5'-二磷酸鸟嘌呤三丁基盐盐,Zncl 2 / dmf,40°C,18 h;对于7m gppporn:5'-二磷酸n 7-甲基鸟苷三丁基盐盐,Zncl 2 / dmf,40°C,18 h; (E)18或19,Cuso 4,抗坏血酸钠,二恶英/H 2 O,65°C MW,1 h; (f)1M DBU / MECN,25°C,3分钟; (g)对于5'OH ORN-X或GPPPORN-X:aq.nh 3,40°C,3 h;对于7m gppporn-X:7M NH 3 / MEOH。

RNA 生物合成和 RNA 代谢因子作为 R 环抑制因子:在基因组完整性中的隐藏作用
DNA 代谢和 RNA 代谢以前被认为是两个不同的研究领域,几乎没有联系。只有现象学观察表明,DNA 序列的转录会增加细菌和酵母中的突变和重组频率(Voelkel-Meiman 等人,1987 年;Dul 和 Drexler,1988 年;Stewart 和 Roeder,1989 年;Thomas 和 Rothstein,1989 年;Beletskii 和 Bhagwat,1996 年)。20 世纪 70 年代和 80 年代对噬菌体、细菌和酵母的遗传分析表明,转录增强了 DNA 受损的倾向,但其分子基础尚不清楚(有关综述,请参阅 Aguilera,2002 年)。然而,随着世纪之交,对导致超重组或超突变的新条件的不同研究揭示了 RNA 代谢与 DNA 完整性之间以前未预见到的联系。这为理解转录相关突变 (TAM) 和转录相关重组 (TAR) 等现象带来了新的视角(有关综述,请参阅 Aguilera 和 Gómez-González 2008)。

实现特定 RNA 靶向和降解的新型化学方法:应用于 SARS-CoV-2 RNA
当前药物化学面临的主要挑战之一是使用小分子药物靶向非编码 RNA。与传统的药物发现方法不同,它需要改变视角才能取得成功。正如 Disney 及其同事 1 在本期《ACS Central Science》中所描述的那样,针对 SARS-CoV-2 RNA 的特异性配体的发现、核糖核酸酶靶向嵌合体 (RIBOTAC) 的开发以及使用尖端化学生物学工具对细胞内作用机制的详细研究,凸显了新化学模式可能对未来疗法产生的影响。这项工作的应用领域涉及针对导致当前全球健康危机的病毒 SARS-CoV-2。人们在药物重新定位、大规模筛选和疫苗开发方面做出了许多努力;然而,显然需要研究用于创新治疗方法的新靶点和新生物活性分子,以增加抗病毒药物库。RNA 被认为是基因表达过程中的中间体,现在是药物发现的有效靶点。 2 RNA 是一种大分子,参与转录、翻译和基因表达调控等重要生物过程。事实上,非编码 RNA 表达和/或功能的许多失调与许多病理直接相关,例如神经系统疾病、心脏病或癌症。最值得注意的是,超过 70% 的人类基因组在非编码 RNA 中转录,而只有 1.5% 编码蛋白质。由于这些蛋白质中只有一小部分代表上市药物的实际靶标,因此很明显,将非编码 RNA 添加到潜在治疗靶标将大大增加药物开发的前景。由新型 RNA 病毒 SARS-CoV-2 引起的冠状病毒大流行凸显了 RNA 靶向疗法在治疗 RNA 病毒引起的感染方面的潜力。这些疗法之前曾在文献中用于 HIV、HCV 或流感病毒。自 20 世纪 40 年代第一批 RNA 配体作为抗生素进入市场以来,新的 RNA 靶向药物的发现取得了重大进展,例如氨基糖苷类或四环素类,以及最近的恶唑烷酮类。3 近年来,RNA 结合剂的进展包括不同类型的原创、有时不寻常的药物发现策略。还开发了用于识别非常特异性 RNA 配体的先导识别策略,例如 Inforna。4 后者是一种基于二维组合筛选 (2-DCS) 和通过测序 (StARTS) 实现的结构-活性关系相结合的技术。将这种统计方法应用于筛选结果可以预测 RNA 库成员的亲和力和选择性并评分结合相互作用。4 该方法已被证明非常有效,可以成功预测特定 RNA 二级结构的化合物。各种筛选技术(例如微阵列或基于荧光的检测)也已用于发现 RNA 配体以及基于结构的选择性配体设计。5-7

RNA 生物合成和 RNA 代谢因子作为 R 环抑制因子:在基因组完整性中的隐藏作用
DNA 代谢和 RNA 代谢以前被认为是两个不同的研究领域,几乎没有联系。只有现象学观察表明,DNA 序列的转录会增加细菌和酵母中的突变和重组频率(Voelkel-Meiman 等人,1987 年;Dul 和 Drexler,1988 年;Stewart 和 Roeder,1989 年;Thomas 和 Rothstein,1989 年;Beletskii 和 Bhagwat,1996 年)。20 世纪 70 年代和 80 年代对噬菌体、细菌和酵母的遗传分析表明,转录增强了 DNA 受损的倾向,但其分子基础尚不清楚(有关综述,请参阅 Aguilera,2002 年)。然而,随着世纪之交,对导致超重组或超突变的新条件的不同研究揭示了 RNA 代谢与 DNA 完整性之间以前未预见到的联系。这为理解转录相关突变 (TAM) 和转录相关重组 (TAR) 等现象带来了新的视角(有关综述,请参阅 Aguilera 和 Gómez-González 2008)。