XiaoMi-AI文件搜索系统
World File Search SystemRNAi
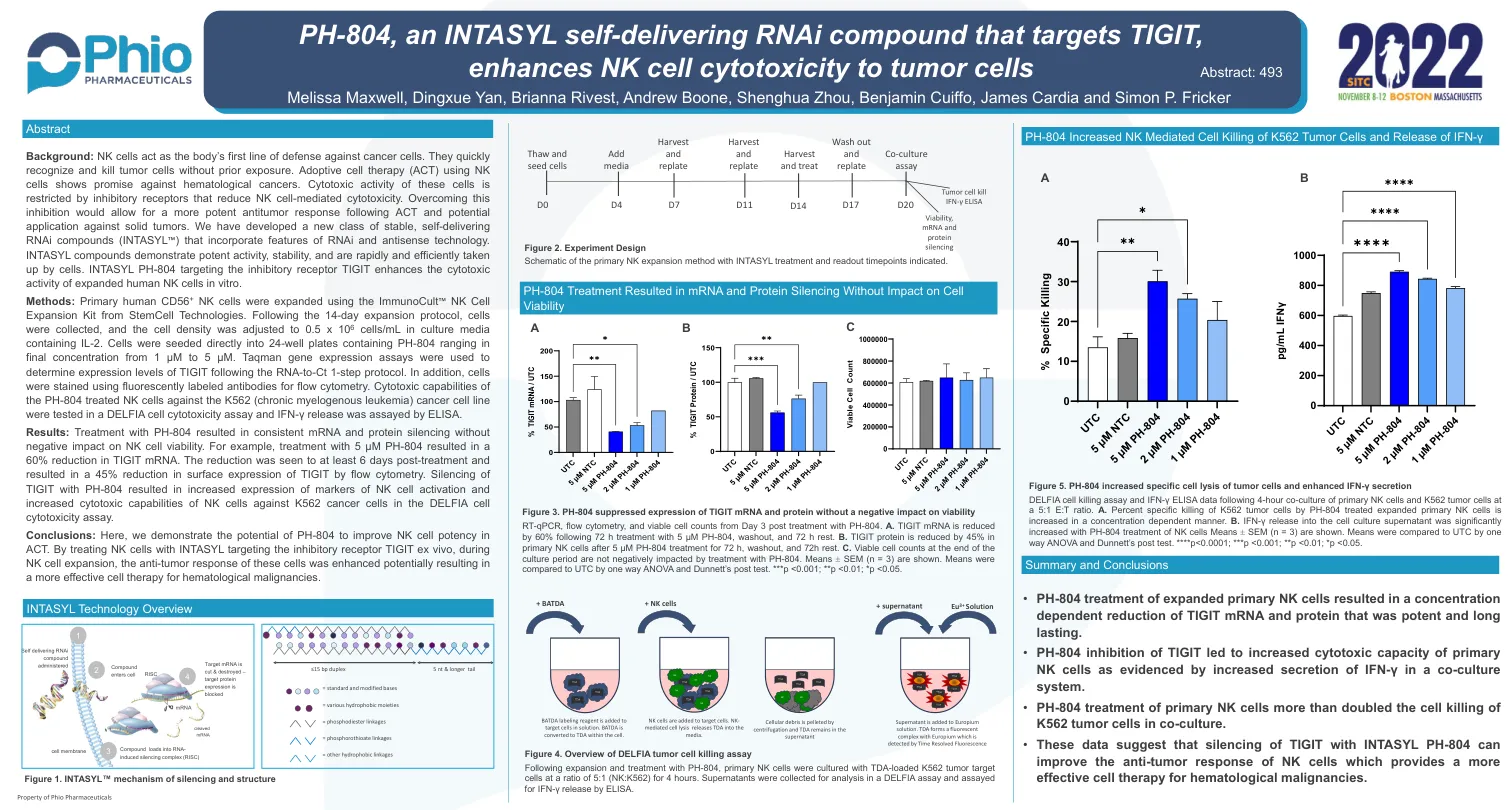
ph-804,一种intasyl自降低的RNAi化合物...
背景:NK细胞充当人体对癌细胞的第一道防线。他们迅速识别并杀死肿瘤细胞而无需事先暴露。使用NK细胞的产卵细胞疗法(ACT)显示出对血液癌的有望。这些细胞的细胞毒性活性受抑制受体的限制,这些受体降低了NK细胞介导的细胞毒性。克服这种抑制作用将允许在ACT和对实体瘤的潜在应用后产生更有效的抗肿瘤反应。,我们开发了一种新的稳定,自我保留的RNAi化合物(Intasyl™),该化合物(Intasyl™)结合了RNAi和反义技术的特征。intasyl化合物表现出有效的活性,稳定性,并且被细胞迅速有效地吸收。靶向抑制受体TigIT的内烷基pH-804在体外增强了扩展的人NK细胞的细胞毒性活性。

口服RNAi用于蚊子中的基因沉默 弥合母亲培养皿中的差距 Xu,D.,Zhou,D.,Bum-erdene,K.,Bailey,B.J.,Sishtla,K.,Liu,S.,Wan,J.,Aryal,U.K. 通过ASD中的模仿学习熟练运动 评估温带森林的生长和气候灵敏度,以响应长期全水水酸化实验 使用横断范式增强维修... 与插管儿科患者相关的呼吸道病原体 中枢神经系统的嘌呤能受体
媒介蚊子传播各种医学上重要的致病病原体(疾病控制中心2021)。矢量控制是预防人类蚊子传播疾病的主要方法。然而,由于杀虫剂抗性的全球发病率不断增加,并担心化学农药对非目标生物的潜在负面影响,当前的蚊子控制方法达到了可持续性的局限性,需要开发和引入创新的矢量控制策略(AIRS和BartholoMay 2017,疾病控制疾病,对疾病控制20221)。蚊子基因组项目(Holt等人2002,Nene等。 2007)促进了蚊子生物学新方面的研究,包括医学上重要的艾园(登革热,Zika,chikungunya和黄热病载体)的功能性遗传研究,以及肛门(疟疾载体)人类疾病媒介(疾病控制中心2021)。 这些进步加剧了以基因为中心的新型载体控制策略的发展,导致研究的研究重点是鉴定潜在的基因靶向载体控制基因靶标,以及操纵蚊子基因在实验室中以及在现场中的作用的方法。 RNAi,促进实验室中蚊子基因的功能表征,2002,Nene等。2007)促进了蚊子生物学新方面的研究,包括医学上重要的艾园(登革热,Zika,chikungunya和黄热病载体)的功能性遗传研究,以及肛门(疟疾载体)人类疾病媒介(疾病控制中心2021)。这些进步加剧了以基因为中心的新型载体控制策略的发展,导致研究的研究重点是鉴定潜在的基因靶向载体控制基因靶标,以及操纵蚊子基因在实验室中以及在现场中的作用的方法。RNAi,促进实验室中蚊子基因的功能表征,

识别活性 RNAi 通路
+HUHZHSUHVHQWWZRVLJQ 4QGLQJVWKDWFRQWULEXWH XQGHUVWDQGLQJRI 白色念珠菌DOLIH-WKUHDWHQLQJ KXPDQIXQJDOSDWKRJHQ)LU HVWDEOLVKWKDWWKH 白色念珠菌UHIHUHQFHVWUDLQLVGHIHF 51$LQWHUIHUHQFHDIXQGDP UHJXODWRU\SDWKZD\6HFRQ GLVFRYHUWKDWLQFRQWUD UHIHUHQFHVWUDLQWKHYD PDMRULW\RIC。白色念珠菌LVRODWH FRQWDLQDQDFWLYH51$ LQWHUIHUHQFH51$ LSDWKZ VLOHQFHVJHQHH[SUHVVLRQ &RQVLGHULQJWKDW51$ LSOD FHQWUDOUROHVLQUHYHUVL JRYHUQLQJJHQHH[SUHVVLRQ JHQRPHVWDELOLW\GUXJUH DQGFRXQWHULQJYLUDOLQIH RXU4QGLQJRHUVYDOXDEOH LQVLJKWVLQWRWKHELRORJ GDQJHURXVIXQJDOSDWKRJHQ

flyrnai.org - 果蝇RNAi筛查中心和转基因RNAi项目的数据库:2021 Update
果蝇RNAi筛查中心和转基因RNAi项目(DRSC / TRIP)的Flyrnai数据库提供了一系列在线资源,可促进功能基因组学研究,并特别强调果蝇Melanogast。目前,数据库提供:以基因为中心的源,可促进直系同源映射和刻度有关通用基因模型物种中直系同源物的信息;以试剂为中心的资源,可帮助研究人员识别RNAi和CRISPR SGRNA试剂或设计;以及以数据为中心的资源,以促进转录数据的可视化和挖掘,蛋白质修饰数据,蛋白质间相互作用等。在这里,我们讨论了有助于生物学和生物医学搜索者有效识别,可视化,分析和整合果蝇和其他物种的信息和数据的更新和新功能。,这些资源共同促进了功能基因组学的多个步骤,从构建基因和试剂列表到数据的管理,分析和集成。

IPLANTA Webinar 9:RNAi植物表观遗传学和发展IPLANTA Webinar 9:RNAi植物表观遗传学和发展
pols = RNA聚合酶; SHH1 = Sawadee同源域同源物1; RDRS = RNA定向的RNA聚合酶; clsy1 =经典1; dcl3 = dicer样3; Hen1 = Hua增强剂1; Ago4 = Argonaute 4; ktf1 =含KOW域的转录因子1; RDM1 = RNA指导的DNA甲基化1; drm2 =域重新排列的甲基转移酶2; DRD1 = RNA导向中有缺陷; DNA甲基化1; dms3 =分生组织沉默3; MORC6 = Microdorchidia 6; idn2 =参与从头2; HDA6 =组蛋白脱乙酰基酶6; JMJ14 = Jumonji 14; ubp26 =泛素特异性蛋白酶26

Zodasiran,RNAi治疗性靶向Angptl3,用于...
来自新陈代谢和脂质保护,纽约西奈山的西奈山脉心脏医院,伊坎医学院(R.S.R.);蒙特利尔大学(D.G.)和伦敦的Robarts Research Institute(R.A.H.)- 两者在加拿大;贝勒医学院和德克萨斯心脏研究所 - 都在休斯敦(C.M.B.);维多利亚大学莫纳什大学莫纳什维多利亚州心脏研究所(S.J.N.)和西澳大利亚大学医学院和珀斯皇家珀斯医院心脏病学系(G.F.W.)- 全部在澳大利亚;卢卡斯研究,北卡罗来纳州莫尔黑德市(K.J.L.);和加利福尼亚州帕萨迪纳(J.S.M.,R.Z.,M.M.,T.C.,J.H。)和Arrowhead Pharma-Ceuticals。可以通过Robert .Rosenson@MSSM .EDU与Rosenson博士联系,或者可以在Sinai Mount Sinai Sinai Medical of Sinai的Sinai fuster Heart Hospital,Sinai Mount Sinai,1 Gustave L. Levy Pl。

RNA 干扰 (RNAi) 和药物的未来...
• 2001 年,研究人员证实 siRNA 介导的基因沉默确实发生在人类细胞中。3 这种形式的基因沉默后来被广泛称为 RNA 干扰,简称 RNAi。2006 年,Andrew Z. Fire 博士和 Craig C. Mello 博士因发现控制遗传信息流动的基本 RNA 机制而获得诺贝尔医学奖。4 • 2018 年,经过多年研究,ONPATTRO® (patisiran) 获得美国食品药品监督管理局 (FDA) 批准用于治疗成人遗传性转甲状腺素蛋白介导 (hATTR) 淀粉样变性多发性神经病,标志着一类全新药物的到来。使用相同的 RNAi 技术,GIVLAARI®(givosiran)于 2019 年获批用于治疗急性肝卟啉症 (AHP) 成人患者,OXLUMO®(lumasiran)于 2020 年获批用于治疗原发性高草酸尿症 1 型 (PH1),以降低儿童和成人患者的尿液草酸水平,Leqvio®(inclisiran)* 于 2021 年获批用于治疗高胆固醇血症。

洞察癌症RNAi治疗的前景
RNA干扰(RNAi)也称为基因沉默,是一种阻止某些疾病(例如癌症)中基因表达的生物过程。它可用于提高治疗(尤其是基因疗法)的准确性、效率和稳定性。然而,面临着诸如将寡核苷酸药物输送到身体较难接触的部位以及毒副作用发生率高等挑战。因此,必须改善它们向目标部位的输送,并减少它们对非癌细胞的有害影响,以充分发挥它们的潜力。在本研究中,讨论了RNAi在治疗困扰许多国家的新型冠状病毒疾病COVID-19中的作用。本综述旨在确定RNAi的机制和应用,并通过确定一些癌症输送系统并提供药物信息以改进它们,探讨RNAi治疗的当前挑战。值得一提的是,脂质基输送系统和外泌体等输送系统通过降低其免疫原性和提高其细胞亲和力,彻底改变了RNAi疗法。更深入地了解 RNAi 在癌症治疗中的机制和挑战可以为 RNAi 药物开发提供新的见解。

体内 RNAi:生物分布、递送和应用。
图 1:100% 人血清中未修饰和 siSTABLE v2 修饰 siRNA 的稳定性。未修饰 siRNA 几乎立即降解,而 siSTABLE siRNA 可保持完整性长达 5 天。

鉴定白色念珠菌中的活性 RNAi 通路
RNA 干扰 (RNAi) 是一种基本调控途径,具有广泛的功能,包括调节基因表达和维持基因组稳定性。尽管 RNAi 在真菌界广泛存在,但众所周知的物种,如模型酵母酿酒酵母,已经失去了 RNAi 途径。到目前为止,还没有证据表明白色念珠菌中存在完全功能的 RNAi 途径,白色念珠菌是一种被世界卫生组织认为至关重要的人类真菌病原体。在这里,我们证明了广泛使用的白色念珠菌参考菌株 (SC5314) 在编码中心 RNAi 成分 Argonaute 的基因中含有失活错义突变。相比之下,大多数其他白色念珠菌分离株含有典型的 Argonaute 蛋白,预计该蛋白具有功能性和 RNAi 活性。事实上,使用高通量小RNA和长RNA测序结合无缝CRISPR/Cas9基因编辑,我们证明了活性白色念珠菌RNAi机制抑制了亚端粒基因家族的表达。因此,白色念珠菌中存在完整且功能性的RNAi通路,这凸显了在研究这种危险病原体时使用多种参考菌株的重要性。