XiaoMi-AI文件搜索系统
World File Search SystemRaza

论文:2型糖尿病在儿童和青少年中 div>
简介:糖尿病被认为是全球健康问题。 div>在过去的十年中,已经报道了小儿种群中新的糖尿病2型糖尿病病例的增加。 div>直到8年前,CMN综合医院“ La Raza”的小儿内分泌服务中还没有记录这种糖尿病。 div>目前有71例糖尿病诊断(DM)2型患者。 div>目的:确定DM 2型小儿种群中的clfnic,人体测量和代谢特征。设计:横向,观察性,描述性调查,无方向性。 div>材料和方法:研究了71名儿童和青少年的临床诊断,对男性的DM 2、31和女性的临床诊断,平均年龄为12.8±2.3和13。。分别为7±1 .8。 div>每个人都研究了诊断的心血管危险因素和症状的家族史;进行了人体测量法,并在幼体后相提取静脉血样品,以测定葡萄糖,脂质和脂蛋白。 div>女性比男性的BMI显着,尽管它没有达到统计意义,但与儿童相比,女性的年龄也谨慎。 div>诊断,体重,taifa,腰部和臀部周长,心率和血压的年龄相似。 div>男性的禁食葡萄糖的含义较低。 div>在两个性别中,CT,C-LDL和C-HDL的平均水平相似。 div>甘油三酸酯的水平在女孩中谨慎地更大,尽管没有达到统计意义。 div>在女性中,发现高胆固醇血症,高甘油三酯血症和高血压的较高患病率,而两组中低α-丙型蛋白血症和肥胖的频率相似。 div>在诊断时,两性占主导地位的最重要的临床表现是多毒,多尿,体重减轻,棘皮动物鼻孔,食欲不振和糖尿病性酮症酸中毒。 div>结论该小组的2型糖尿病和肥胖症的阳性家族病史很高。 div>糖尿病儿童诊断出多次多次,多尿,食欲丧失,棘皮动物刺痛和肥胖。 div>这些患者具有不利的脂质特征和血脂异常的高频率。 div>

基林独立学区
•书签项目:在PDF上,右键单击并选择“显示导航窗格按钮”。查找书签的符号,然后单击符号。从那里您可以选择所需的主题。•查找:在导航搜索中输入课程号,课程名称或短语,然后单击“ Enter”。继续单击Enter将转到使用此号码或名称的下一页。本课程选择指南旨在帮助您和您的父母设计适合您需求的教学计划。在与辅导员见面以安排课程之前,请花时间研究课程的描述。已经提供了学生工作表供您使用。本指南可能出于各种原因列出了下一学年不会提供的课程。将在受托人批准后添加任何新课程。有关调度和/或注册的信息,请致电您的辅导员以获取高中校园和地区信息页面上列出的适当号码的指导。注释HNRS荣誉AP高级位置DC双信用先决条件 *先决条件需要成功完成。*荣誉,双重信用和AP已被确定为满足GT学生的需求。2015年基林独立学区不会在其计划和活动中区分种族,颜色,国籍,性别,残疾或年龄。已指定以下人员来处理有关非歧视政策的查询:德克萨斯州基林市北WS Young Drive 200号专业标准管理员。76543,254-336-0000。kisd no Incrimina Contra Raza,颜色,Origen Nacional,Género,Incapacidad o Edad e Sus an Sus y Adividades。la sigunite persona ha sido distionada para para precuntas relacionadas conlapólizadediviminación:行政行政专业人士专业人士200 North WS Young Drive,Killen,TX 76543,76543,254-336-0000。基伦独立学区(Killen独立学区)不允许基于标志的品种,肤色,国籍,蝎子,残疾或年龄的歧视。如果有关于非歧视指令的问题,则可以使用以下人员:专业标准管理员200 North W S Young Drive,Killen,TX 76543,254-336-0000。

副教授教授奥斯曼·通克
副教授 Osman Tunç 个人信息 电子邮件:osmantunc@yyu.edu.tr 网址:https://avesis.yyu.edu.tr/osmantunc 国际研究人员 ID ORCID:0000-0003-2965-4561 ScopusID:56638410400 Yoksis 研究人员 ID:330454 已发表的期刊文章被 SCI、SSCI 和 AHCI 检索 I. 论具有无界和分布延迟以及主要非延迟项的非线性系统的全局稳定性 Braverman E.、Tunç C.、Tunç O. 非线性科学与数值模拟通信,第 143 卷,2025 年(SCI 扩展版)II。 Peyrard-Bishop 振荡器链模型中分数 DNA 动力学的分析研究 Riaz MB、Fayyaz M.、Rahman RU、Martinovic J.、Tunç O. Ain Shams 工程杂志,第 15 卷,第 8 期,2024 年 (SCI-Expanded) III. 非线性耦合双曲空间非齐次系统 Lp 范数中的指数稳定性Slynko V.、Tunç O.、Atamas I. 应用数学和计算,第 472 卷,2024 年 (SCI-Expanded) IV。 Caputo 分数阶延迟微分方程的 Ulam–Hyers–Mittag–Leffler 稳定性的新结果 Tunç O.Mathematics,第 12 卷,第 9 期,2024 年 (SCI-Expanded) V. 探索非线性分数 Gilson-Pickering 方程的解析解和调制不稳定性 Rahman RU、Riaz MB、Martinovic J.、Tunç O. Results in Physics,第 57 卷,2024 年 (SCI-Expanded) VI. 具有多重延迟的二阶脉冲微分方程的存在性和稳定性 Pinelas S.、Tunç O.、Korkmaz E.、Tunç C. Electronic Journal of Differential Equations,第 2024 卷,2024 年 (SCI-Expanded) VII.非线性脉冲多重滞后微分和二阶脉冲积分微分方程解的存在性 Bohner M.,Tunç O.,Tunç C. 非线性和凸分析杂志,第 25 卷,第 9 期,第 2337-2360 页,2024 年 (SCI-Expanded) VIII. 迭代积分方程的 ULAM 型稳定性 Tunç O.非线性和凸分析杂志,第 25 卷,第 8 期,第 1899-1908 页,2024 年 (SCI-Expanded) IX.通过森林管理计划减轻大气二氧化碳影响的建模和模拟 Riaz MB、Raza N.、Martinovic J.、Bakar A.、Tunç O. AIMS Mathematics,第 9 卷,第 8 期,第 22712-22742 页,2024 年(SCI-Expanded)

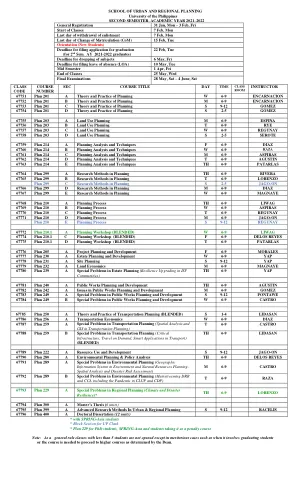
城市与区域规划学院
67751 计划 201 A 规划理论与实践 W 6-9 ENCARNACION 67752 计划 201 B 规划理论与实践 M 6-9 ENCARNACION 67753 计划 201 C 规划理论与实践 S 9-12 GOMEZ 67754 计划 201 D 规划理论与实践 S 2-5 GOMEZ 67755 计划 203 A 土地利用规划 M 6-9 ESPINA 67756 计划 203 B 土地利用规划 T 6-9 RYE 67757 计划 203 C 土地利用规划 W 6-9 REGUNAY 67758 计划 203 D 土地利用规划 S 2-5 SEROTE 67759 计划 214 A 规划分析与技术 F 6-9 DIAZ 67760 计划 214 B 规划分析与技术 W 6-9 RAZA 67761 计划 214 C 规划分析与技术 M 6-9 ASPIRAS 67762 计划 214 D 规划分析与技术 T 6-9 AGUSTIN 67763 计划 214 E 规划分析与技术 TH 6-9 PATARLAS 67764 计划 299 A 规划研究方法 TH 6-9 RIVERA 67765 计划 299 B 规划研究方法 T 6-9 LORENZO 计划 299 C 规划研究方法 S 2-5 JAGO-ON 67766 计划 299 D 规划研究方法 M 6-9 DIAZ 67767 计划 299 E 规划研究方法 W 6-9 MAGNAYE 67768 计划 210 A 规划过程 TH 6-9 LIWAG 67769 计划 210 B 规划过程 W 6-9 ASPIRAS 67770 计划 210 C 规划过程 T 6-9 REGUNAY 67771 计划 210 D 规划过程 M 6-9 JAGO-ON 计划 210 E 规划过程 S 9-12 REGUNAY 67772 计划 210.1 A 规划研讨会(混合) W 6-9 LIWAG 67774 计划 210.1 C 规划研讨会(混合) F 6-9 DELOS REYES 67775 计划 210.1 D 规划研讨会(混合) T 6-9 PATARLAS 67776 计划 205 A 项目规划与开发 F 6-9 MORALES 67777 计划 230 A 房地产规划与开发 W 6-9 YAP 67778 计划 231 A 场地规划 S 9-12 YAP 67779 计划 232 A 土地经济学 M 6-9 MAGNAYE 67780 计划 239 A 遗产规划中的特殊问题(ISF 社区的复原力升级)

微生物取证和计算生物学
[19] Kunin,V.,Copeland,A.,Lapidus,A.,Mavromatis,K。,&Hugenholtz,P。(2008)。宏基因组学的生物信息学指南。微生物学和分子生物学评论,72(4),557-578。[20] Jolley,K。A.,Chan,M。S.,&Maiden,M.C。(2004)。MLSTDBNET分布的多洛克斯序列键入(MLST)数据库。BMC生物信息学,5(1),86。[21] Enright,M。C.和Spratt,B。G.(1999)。多焦点序列键入。微生物学的趋势,7(12),482-487。[22] Healy,M.,Huong,J.,Bittner,T.,Lising,M.,Frye,S.,Raza,S。,&Woods,C。(2005)。通过自动重复序列的PCR键入微生物DNA。临床微生物学杂志,第43(1)期,199-207。[23] Vergnaud,G。和Pourcel,C。(2006)。多个基因座VNTR(串联重复的可变数量)分析。分子鉴定,系统学和原核生物的种群结构,83-104。[24] Van Belkum,A。(2007)。通过多焦点数量的串联重复分析(MLVA)来追踪细菌物种的分离株。病原体和疾病,49(1),22-27。[25] Vergnaud,G。和Pourcel,C。(2009)。多个基因座变量串联重复分析数。微生物的分子流行病学:方法和方案,141-158。[26] Fricke,W。F.,Rasko,D。A.和Ravel,J。(2009)。基因组学在鉴定,预测和预防生物学威胁中的作用。PLOS Biology,7(10),E1000217。[27] Wu,M。和Eisen,J。A.(2008)。95-100)。一种简单,快速且准确的系统基因推断方法。基因组生物学,9(10),R151。[28] Liu,B.,Gibbons,T.,Ghodsi,M。和Pop,M。(2010年12月)。隐式:元基因组序列的分类分析。生物信息学和生物医学(BIBM),2010年IEEE国际会议(pp。IEEE。 [29] Wang,Z。,&Wu,M。(2013)。 门水平细菌系统发育标记数据库。 分子生物学与进化,30(6),1258-1262。 [30] Darling,A。E.,Jospin,G.,Lowe,E.,Matsen IV,F。A.,Bik,H。M.,&Eisen,J. A. (2014)。 系统缩影:基因组和宏基因组的系统发育分析。 peerj,2,e243。 [31] Taberlet,P.,Prud'Homme,S.M.,Campione,E.,Roy,J.,Miquel,C.,Shehzad,W。,&Melodelima,C。(2012)。 土壤采样和细胞外DNA的分离,适用于大量的起始材料。 分子生态学,21(8),1816-1820。IEEE。[29] Wang,Z。,&Wu,M。(2013)。门水平细菌系统发育标记数据库。分子生物学与进化,30(6),1258-1262。[30] Darling,A。E.,Jospin,G.,Lowe,E.,Matsen IV,F。A.,Bik,H。M.,&Eisen,J.A.(2014)。系统缩影:基因组和宏基因组的系统发育分析。peerj,2,e243。[31] Taberlet,P.,Prud'Homme,S.M.,Campione,E.,Roy,J.,Miquel,C.,Shehzad,W。,&Melodelima,C。(2012)。土壤采样和细胞外DNA的分离,适用于大量的起始材料。分子生态学,21(8),1816-1820。

支持 MEC 的 IoV 网络中 AI 赋能的数据卸载
Error 500 (Server Error)!!1500.That’s an error.There was an error. Please try again later.That’s all we know.Error 500 (Server Error)!!1500.That’s an error.There was an error. Please try again later.That’s all we know.


拉斯孔德斯诊所医学杂志
摘要 科学的进步表明,在不久的将来,通过作用于生殖细胞或植入前胚胎来修改新个体的基因的可能性将在整个人口中实现。 2018年底,国际科学界对贺建奎博士的实验表示担忧。贺建奎利用CRISPR-Cas 9技术,对人类胚胎进行生殖目的的基因改造,导致至少两名女婴诞生。在本文中,我们将按照伊曼纽尔提出的标准,对建奎博士的实验进行伦理分析;根据这种观点,该实验不符合科学伦理委员会在评估方案时通常使用的任何道德标准。然后,我们将回顾有关在人类生殖细胞(精子和卵子)和植入前胚胎中使用基因编辑用于生殖目的的伦理争议。由于这些变化可以遗传给后代,而且该技术仍处于实验阶段,我们将主张暂停将其用于这些目的。当基因编辑不是用于生殖目的,而是用于产生新知识时,我们将解释为什么我们认为有必要区分该技术在生殖细胞中的应用与人类胚胎的研究,这一区别可能会根据对人类胚胎作为人类物种活体的评估而受到质疑。我们还将简要讨论使用基因编辑技术治疗或预防疾病与使用基因编辑技术来改善或“增强”人类之间的区别,因为后者存在一些道德上的异议。最后,我们将对智利和国际上的监管观点进行简要分析,因为这些技术的一些应用引发了道德问题,凸显了对该领域进行严格监管的必要性。

植物育种技术的演变
D Shashibhushan 和 Ashish Reddy Muchanthula 摘要 植物育种是一门改变植物性状以产生所需特性的科学。为了改善与作物各种性状相关的农艺性状,已经使用了几种常规和分子方法,包括遗传选择、基于全基因组序列的方法、物理图谱和功能基因组工具。然而,使用可编程核酸酶和 CRISPR 相关 (Cas) 蛋白的基因组编辑技术的最新进展为新的植物育种时代打开了大门。因此,为了提高作物育种的效率,世界各地的研究人员正在使用新策略,例如快速育种、基因组编辑工具和高通量表型分析。在这篇综述中,我们总结了作物育种几个方面的最新发现,以描述植物育种实践从传统到现代快速育种的演变。 关键词:脱氧核糖核酸 (DNA)、核糖核酸 (RNA) CRISPR、基因沉默、基因组编辑、反向育种 简介 农业始于大约 10,000 年前。从那时起,人类在不知不觉中就选择植物来满足自己的需求。首先,只有自然界提供的性能最好的植物才会被选择和保留。自发出现的有用特性通过人类选择培育成某些作物,通常是违背自然选择的;因此,在没有任何科学方法的情况下进行植物育种。当时孟德尔遗传定律的知识还不为人所知。19 世纪末,孟德尔定律被发现,这加速了植物改良。1953 年,沃森和克里克提出了 DNA 双螺旋模型,大大增加了人们对遗传物质的理解。这是植物育种的一个重大转折,因为针对 DNA 的植物改良开始曝光,第一个是 20 世纪 60 年代的突变育种,后来是 20 世纪 80 年代的转基因技术。从那时起,遗传学科学从不同的 DNA 分析方法到标记辅助选择,突飞猛进。虽然已经发现了许多不同的技术,但它们仍然是独一无二的,每种技术都适用于特定情况。多种技术的出现为植物育种者提供了培育新品种所需的“工具”。为什么这是一个永无止境的过程?“植物育种是一个连续的过程”。这句话自古以来就没有过时。为了满足消费者的需求,植物育种在粮食安全和食品安全中发挥着重要作用。然而,由于人口的急剧增长,植物育种在全球范围内面临着食品质量和数量的问题。在这个快节奏的时代,消费者更喜欢即食食品,而营养质量却有所下降。此外,气候变化导致的天气条件变化正在导致高温和干旱胁迫;因此,世界各地的农民都面临着严重的产量损失。预计到 2050 年,世界人口将达到 100 亿。考虑到这一点,必须在有限的土地上利用有限的资源培育新品种。古老的植物育种实践虽然没有失去其重要性,但仅靠这些还不足以满足当前的粮食需求状况 (Raza et al ., 2019) [21] 。此外,植物育种也面临着自身的挑战。它的作用是创造新的等位基因组合,固定所需的等位基因并控制基因流动。考虑到上述标准,植物育种应该是一个永恒的关注和进步的主题。植物育种,从传统方法到如今与现代生物技术工具的结合,在过去几年中发展迅速。随着时间的推移,人们在为不同目的培育植物方面取得了许多进步。每一项进步,