XiaoMi-AI文件搜索系统
World File Search SystemS6K

通过HEY1和ID2B信号传导调节MüllerGlia对视网膜损伤的再生反应
标准授予问题1绑定RET [M918T]] 5绑定BRAF,SRC,S6K 1、3或9如果这些目标的1、2或3绑定避免避免MKNK1* 3避免TTK,ERK8,PDK,PDK,PDK,PAK3 1、2、3或4如果它避免了AR bak,则避免了这些目标,如果它避免了5个目标。结合FGFR1,LKB 1或3,如果这些靶标的结合1或2避免PAK3* 3避免MAP3K7* 3避免了PIK3CA 1进入CNS 3的PIK3CA 1 3这两个问题新颖的能力2使用专利2类药物3类药物3表1-对问题1和2。标明目标表明182

MicroRNA-126-3p 抑制 HeLa 细胞增殖,...
摘要。我们之前曾报道,与正常宫颈粘液相比,microRNA 126-3p (miR-126-3p) 在患有明显宫颈癌或癌前病变的患者的宫颈粘液中的含量明显更高。在本文中,我们研究了在宫颈癌细胞系 HeLa 中强制表达 miR-126-3p 对增殖、迁移、侵袭、凋亡和蛋白质表达的影响。我们用 miR-126-3p miRNA 转染 HeLa 细胞,发现这些细胞的增殖、迁移和侵袭(通过细胞计数、伤口愈合、细胞迁移和侵袭测定)相对于用阴性对照模拟物转染的细胞显著降低。在 miR-126-3p 转染的细胞中,磷酸肌醇 3 激酶 (PI3K)、磷酸化 3-磷酸肌醇依赖性蛋白激酶-1 (p-PDK1) 和 p-AKT 蛋白的水平较低。磷酸化 70S6K (p-p70S6K)、磷酸化糖原合酶激酶 3 β (p-GSK3 β )、磷酸化 S6K (p-S6K)、细胞周期蛋白 D1、磷酸化 p21 活化激酶 1 (p-PAK1)、Rho 相关卷曲螺旋蛋白激酶 1 (ROCK1)、肌强直性营养不良相关 CDC42 结合激酶 α (MRCK α ) 和磷脂酶 C γ 1 (p-PLC γ 1) 也下调。这表明 PI3K/PDK1/AKT 通路的下游效应子是 miR-126-3p 抑制的靶标。相反,凋亡相关蛋白,包括 BCL-2 相关细胞死亡激动剂 (Bad)、B 细胞淋巴瘤特大 (Bcl-xL) 和 BCL-2 相关 X (Bax),均被 miR-126-3p 上调,导致 caspase 3/7 活性增加和细胞凋亡。因此,miR-126-3p 的强制表达

抗 CD25 靶向疗法作为免疫调节剂和癌症靶向药物的新兴作用
图 1. 受刺激 T 细胞中的 IL2R 激活途径表示。IL2R 的不同构象会影响其对 IL2 的亲和力(低亲和力 CD25 或高亲和力三聚体受体)。IL2 还可以通过 CD122/CD132 二聚体影响信号传导。此外,形成受体的 CD25 分子是来自相邻细胞(反式)还是同一细胞(顺式)决定了高亲和力异三聚体受体的命名惯例。途径的激活由 Janus 激酶 1 和 3(JAK1 和 JAK3)磷酸化启动,进而刺激 STAT5 二聚化,或磷酸肌醇 3 激酶 (PI3K) 和大鼠肉瘤病毒致癌基因同源物 (Ras) 途径,最终磷酸化效应激酶 p70 S6K 和 MAPK。
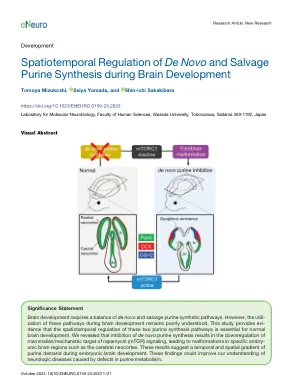
大脑发育过程中从头和挽救嘌呤合成的时空调节
嘌呤的水平,维持真核细胞体内平衡的必需分子,受到从头和救助合成途径的坐标的调节。在胚胎中枢神经系统(CNS)中,从头途径对于满足神经茎/促生细胞(NSPC)主动扩散的要求被认为至关重要。但是,在中枢神经系统开发期间,这两种途径如何平衡或分别使用。在这项研究中,我们显示了途径利用率的动态变化,并且在胚胎阶段和产后 - 成年小鼠脑的拯救途径上更依赖于从头途径。各种嘌呤合成抑制剂在体外的药理作用以及嘌呤合成酶的表达概况表明,胚胎大脑中的NSPC主要使用从头途径。同时,小脑中的NSPC同时需要从头和打捞途径。在从头抑制剂的体内给药导致前脑皮质区域严重下降症,表明沿胚胎大脑的前后轴沿着嘌呤的嘌呤需求梯度,而背侧前脑的皮质区域比腹膜或腹膜较高的嘌呤需求更高。 这种新皮层的组织学缺陷伴随着雷帕霉素复合物1(MTORC1)/核糖体蛋白S6激酶(S6K)/S6信号传导壳的强烈下调,这是一种至关重要的途径,用于细胞代谢,生长和生存。在从头抑制剂的体内给药导致前脑皮质区域严重下降症,表明沿胚胎大脑的前后轴沿着嘌呤的嘌呤需求梯度,而背侧前脑的皮质区域比腹膜或腹膜较高的嘌呤需求更高。这种新皮层的组织学缺陷伴随着雷帕霉素复合物1(MTORC1)/核糖体蛋白S6激酶(S6K)/S6信号传导壳的强烈下调,这是一种至关重要的途径,用于细胞代谢,生长和生存。这些发现表明,嘌呤途径对MTORC1信号传导和适当脑发育的时空调节的重要性。

MAPK和PI3K途径的四倍编辑有效地阻止Kras突变的结直肠癌细胞的进展
包含完整的蛋白酶抑制剂鸡尾酒(Roche,巴塞尔,瑞士)。将提取物通过十二烷基硫酸钠 - 聚丙烯酰胺凝胶电泳(SDS-PAGE)分离,并转移到聚偏二氟化物(PVDF)膜上。膜用5%的非脂肪牛奶或5%牛血清白蛋白(BSA)封闭,然后与针对以下抗原的主要抗体孵育:KRAS:KRAS(#12063-1; Proteintech,Rosemont,Rosemont,IL,IL,USA),PIK3CA,PIK3CA(PIK3CA),PIK3CA(#4249; Celling Signaling Technology; Celling Signaling Technology,Signal,Danvers,Danvers,Ma),Ma,Ma),MA,AKT(29),#29; phospho-AKT (Ser473; #4060; Cell Signaling Technology), MTOR (#2983; Cell Signaling Technology), S6K (#9202; Cell Signaling Technology), phospho-S6K (T389; #9205; Cell Signaling Technology), MEK (#9126; Cell Signaling Technology), ERK (#4695; Cell Signaling Technology), phospho-ERK (Thr202/Tyr204;#4370;细胞信号技术),

小细胞肺癌
富含脯氨酸的15(PRR15)是一种主要以其在胎盘发育中的作用而闻名的蛋白质。这项研究研究了非小细胞肺癌(NSCLC)中PRR15的表达,功能明显和基础机制。与正常的肺实质相比,NSCLC组织中的PRR15表达显着升高,其表达较高与不良临床结局相关。单细胞RNA测序确认在恶性肿瘤细胞群中确认PRR15。PRR15的表达在NSCLC组织中升高,来自局部治疗的患者以及一组原发性和已建立的NSCLC细胞。PRR15使用SHRNA或CRISPR/CAS9介导的敲除的耗竭显着抑制了增殖和迁移,同时促进了各种NSCLC细胞的凋亡。相反,使用慢病毒构建体增强细胞增殖和迁移的异位PRR15过表达。机械研究涉及PRR15在Akt-MTOR信号通路的激活中。通过SHRNA或CRISPR/CAS9介导的敲除对PRR15表达的抑制导致AKT和S6K磷酸化降低,而PRR15过表达导致原代人NSCLC细胞中的Akt-S6K信号扩展。 使用异种移植模型的体内研究进一步验证了PRR15的致癌作用,这表明PRR15敲低抑制了肿瘤的生长并减弱了Akt-MTOR激活。 这些发现共同强调了PRR15作为NSCLC中新型的致癌驱动力和治疗靶标的潜力。通过SHRNA或CRISPR/CAS9介导的敲除对PRR15表达的抑制导致AKT和S6K磷酸化降低,而PRR15过表达导致原代人NSCLC细胞中的Akt-S6K信号扩展。使用异种移植模型的体内研究进一步验证了PRR15的致癌作用,这表明PRR15敲低抑制了肿瘤的生长并减弱了Akt-MTOR激活。 这些发现共同强调了PRR15作为NSCLC中新型的致癌驱动力和治疗靶标的潜力。使用异种移植模型的体内研究进一步验证了PRR15的致癌作用,这表明PRR15敲低抑制了肿瘤的生长并减弱了Akt-MTOR激活。这些发现共同强调了PRR15作为NSCLC中新型的致癌驱动力和治疗靶标的潜力。

双囊MTORC1抑制剂在肿瘤模型中诱导凋亡细胞死亡,以多活化的MTORC1
简介PI3K/AKT/MTOR网络是一种关键的细胞内信号,该途径指导生理和病理条件下的细胞生长和代谢(1)。雷帕霉素(MTOR)进化保守的哺乳动物靶标是在哺乳动物细胞中表达的丝氨酸/苏氨酸激酶(2)。mTOR是MTORC1和MTORC2蛋白复合物中的关键蛋白(3),MTORC1调节参与蛋白质合成,基因表达,葡萄糖和脂质代谢和核苷酸生物合成的信号传导级联反应(4)。mTORC1磷酸化4E结合蛋白1(4EBP1)和S6激酶(S6K),涉及cap依赖性翻译起始和伸长的下游靶标(5)。在结节性硬化症复合物(TSC)肿瘤和多种癌症类型中发生的TSC1或TSC2的完全丧失(6-8)导致MTORC1的组成型非调节激活(9)。鉴于人类肿瘤中PI3K/AKT/MTOR信号的频繁激活,已经开发了几代MTOR抑制剂(1)。Rapalogs治疗了几种与TSC相关的肿瘤以及肾细胞癌(RCC)(10)。但是,旋转在治疗RCC,膀胱癌(BLCA),

研究论文比较神经母细胞瘤细胞中 RIST 分子靶向方案中的 mTOR 抑制剂雷帕霉素和 Torin-2
复发或难治性高危神经母细胞瘤患者的预后仍然不容乐观,迫切需要新的治疗方案。RIST 治疗方案采用多模式节拍疗法设计,结合分子靶向药物(雷帕霉素和达沙替尼)和化疗主干(伊立替康和替莫唑胺),目前正在 II 期临床试验(NCT01467986)中验证。随着新型、更有效的 ATP 竞争性 mTOR 抑制剂的出现,我们期望改进 RIST 联合疗法。通过比较 Torin-1、Torin-2、AZD3147 和 PP242 的 IC 50 值,我们确定只有 Torin-2 抑制了在纳摩尔浓度下测试的所有三种 MycN 扩增神经母细胞瘤细胞系的细胞活力。两种 mTOR 抑制剂的单独治疗诱导了显著的 G 1 细胞周期停滞,与达沙替尼联合治疗降低了细胞周期调节剂细胞周期蛋白 D1 的表达或增加了细胞周期抑制剂 p21 的表达。两种 mTOR 抑制剂的组合指数表明与达沙替尼具有协同作用。有趣的是,与雷帕霉素相比,与 Torin-2 联合治疗可产生更广泛的 mTOR 通路抑制,这表现为 AKT(Thr308、Ser473)、4E-BP(Ser65)和 S6K(Thr389)磷酸化降低。此外,在改良的多模态 RIST 方案中用 Torin-2 替代雷帕霉素可降低细胞活力并诱导细胞凋亡,尽管所用的 Torin-2 药物浓度明显较低。纳摩尔浓度的功效可显著降低与雷帕霉素相关的不必要的免疫抑制。然而,目前我们不能排除 Torin-2 因其在更复杂系统中的效力而增加毒性的可能性。尽管如此,我们的结果表明,在 RIST 方案中加入 Torin-2 作为雷帕霉素的替代品可能是一种有效的选择,可以在复发或治疗难治性高危神经母细胞瘤的前瞻性临床试验中进行评估。