XiaoMi-AI文件搜索系统
World File Search SystemS7

醛介导的天冬酰胺生物合成抑制对糖尿病和酒精中毒的影响
在HCHO处理的LN-18细胞中进行了分析,揭示了天冬酰胺消耗的证据,尽管效果比HEK293T细胞弱(图S5†)。该观察结果至少部分是由于使用非二元胎牛血清的使用,该胎儿血清含有相对较高的天冬水平。与此相一致,与非二元胎儿牛血清一起孵育的HEK293T细胞未观察到氨基酸的耗竭(图S6和S7†)。半胱氨酸分别与HCHO和ACH反应,分别给出硫脯氨酸和2-甲基噻唑烷-4-羧酸(MTCA)(图1)。9,10,27用HCHO或ACH对HEK293T细胞的处理分别导致硫丙啉和MTCA水平升高(图1b)。在HCHO处理的LN-18细胞中也观察到了硫代丙烯的形成(图S5†)。在血28和人类寄生虫中报道了半胱氨酸和ACH对MTCA的反应; 29这里提供的证据还表明,MTCA可能发生在人类细胞中。半胱氨酸-MGO加合物不是
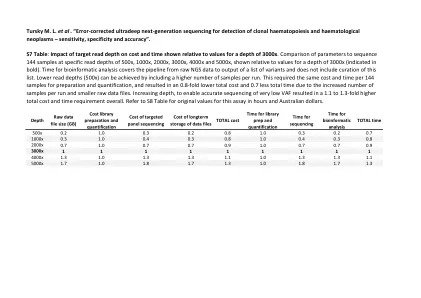
Tursky ML等。 “错误校正的超过下一代...
s7表:目标读取深度对相对于3000倍深度值所显示的成本和时间的影响。在500x,1000x,2000x,3000x,4000x和5000x的特定读取深度上进行参数与序列144个样本的比较,相对于3000x深度的值显示(BOLD表示)。生物信息学分析的时间涵盖了从RAW NGS数据到变体列表的输出的管道,不包括此列表的策划。可以通过在每次运行中包括较大数量的样本来实现较低的读取深度(500倍)。这需要每144个样本的成本和时间进行准备和定量,并且由于每次运行的样本数量增加和较小的原始数据文件,总成本降低了0.8倍,总时间减少了0.7。增加深度,以实现非常低的VAF的准确测序,总体上总成本和时间要求提高了1.1至1.3倍。以小时和澳元的原始值参考S8表。

社会运动和信息城市
6Eg,Alvin Tomer's7カε7巧か4晦re8,对于与此类意识形态预言相矛盾的经验性问答,s㏄:Am Markusen,PeterHAI1,and Amy Glasmeier,疏8hπch加8伽,Londonl AlIen and Unwin,19861 and Thierry Noyelle and Thomas Stanback,7γ1e Eωno'n'c丑oη⑳7η2α!'orπoゾ。4醒ε7'c佣α”θ5,Totowa,N.J.l Allanheld,Osman and Co.,1984, 7Mitchell Moss,“Te1㏄通讯与城市的未来,”in加η41)θりθZop耀π'5'磁85,1986, 3,第33-44页。 8Penny Gurstein,“电子家庭对社会空间模式的影响”,伯克利:加州大学,城市与区域规划系,CP284 研讨会论文,Sp.g1987。 9Margrethe H・01son・“美国在家工作趋势概述”现纽约:纽约大学工商管理研究生院,信息系统研究中心,1983。 10Alelandro Portes,Manuol Castells 和 Lauren Benton(编辑)著,巴尔的摩:约翰霍普金斯大学出版社,1988。

补充材料
正文中显示的计算是使用 Quantum Espresso (QE) 第一性原理程序包 [ S1 , S2 ] 执行的。我们使用密度泛函理论 (DFT) 计算电子结构。使用专为处理表面科学问题而设计的 BEEF-vdW 交换关联函数 [ S3 ]。我们使用 A. dal Corso 的超软伪势 [ S4 , S5 ],动能截止为 1360 eV,电子态占有率的高斯涂抹为 0.27 eV。通过以 Γ 为中心的 12 × 12 × 1 Monkhorst-Pack (MP) 网格 [ S6 ] 对布里渊区进行采样来评估电子态和电荷密度。动力学矩阵和声子微扰势使用 QE 包的 PHonon 代码中实现的密度泛函微扰理论 (DFPT) 进行评估。具体而言,动力学矩阵和微扰势是在 Γ 中心的 6 × 6 × 1 q 网格中进行评估的。我们使用电子声子 Wannier (EPW) 代码来评估电子声子 (e-ph) 矩阵元素 [S7、S8],定义为

mCherry 明亮变体的定向进化:......
图片标题 图 S1 C9 和 C12 变体的 EP-PCR 文库。 图 S2 对 mCherry 家族进行诱变努力以实现寿命进化的总结。 图 S3 空间远距离替换对光物理特性的作用。 图 S4 寿命和酵母细胞共同进化轨迹。 图 S5 哺乳动物细胞的细胞亮度。 图 S6 大肠杆菌的光漂白趋势。 图 S7 比较大肠杆菌在激发速率标准化条件下的光稳定性。 图 S8 归一化的吸收和发射光谱。 图 S9 变体的荧光各向异性衰减和旋转时间常数(τ r)。 图 S10 变体的荧光衰减和平均寿命(τ)。 图 S11 变体的荧光量子产率()。 图 S12 mCherry 变体的辐射速率常数分析。图 S13 总非辐射速率常数与能隙的拟合。图 S14 相关可观测量的示意图。图 S15 波数尺度上的吸光度和荧光光谱。图 S16 方程 S2 中的分子和分母函数。补充数据表列表

最终报告 - SKYbrary
1.1. 飞行历史 ................................................................................................................................................ 11 1.2. 人员受伤 ................................................................................................................................................ 14 1.3. 飞机损坏 ................................................................................................................................................ 14 1.4. 其他损坏 ................................................................................................................................................ 19 1.5. 人员信息 ............................................................................................................................................. 19 1.5.1. 机组人员信息 ................................................................................................................................ 19 1.5.2. 对航空公司波音 737 航班运营的一般性评论 ............................................................................. 28 1.5.3. 俄罗斯联邦“国家空中交通管理公司”塔塔航空导航分公司喀山空中交通管理中心空中交通服务人员的数据 ............................................................................................................................................. 40 1.6.飞机信息................................................................................................................................................ 44 1.6.1. 需要注意的飞机构造特点................................................

突出的细菌分离株用于酿酒厂的脱色水清洗
基于糖蜜的酿酒厂会产生大量的花费,这是一种主要的环境污染物,由于其高的有机负荷和深棕色。这种颜色主要是由黑色素蛋白引起的,黑色素蛋白是通过Maillard反应形成的,Maillard反应是糖和氨基酸之间的非酶促过程。在这项研究中,从40个分离株中选择了八种有希望的细菌菌株,并指定为S1,S2,S3,S4,S5,S5,S6,S7和S8。这些分离株被筛选,以使用定性和定量分析,使酿酒厂消失的洗涤液脱色。中,分离株S5在不同的洗涤浓度(10%,20%和40%)中表现出最高的脱色潜力。值得注意的是,在10%的浓度下,分离株S5完全(100%)脱色,使其成为本研究中最有效的菌株。基于初步表征,分离株S5试初步鉴定为倾斜物种。其特殊的脱色能力表明,它在酿酒厂的生物修复中具有巨大的商业应用潜力。有关优化环境条件并扩大过程的进一步研究,可以为生态友好且具有成本效益的解决方案铺平道路,以减轻酿酒厂废水的环境影响。简介糖蜜酿酒厂是工业污染的主要因素,产生了大量的高强度废水,其生化氧需求(BOD)和化学氧需求(COD)显着升高。这些分离株通过定性和定量分析筛查了消耗清洗的能力。酿酒厂花费的洗涤物中的主要污染物之一是黑色素素,这是一种复杂的化合物,它是通过maillard反应形成的,是糖和氨基酸之间的非酶相互作用。黑色素素特别关注的是,通过减少水体的光渗透,改变微生物生态系统并抑制植物的生长,从而有助于环境降解。[1]在这项研究中,从总共40个分离株中选择了八种有希望的细菌菌株,并指定为S1,S2,S3,S4,S4,S5,S6,S7和S8。中,分离株S5在不同的洗涤浓度(10%,20%和40%)时表现出最高的脱色潜力。值得注意的是,在10%的浓度下,分离株S5在指定时期内达到100%脱色,使其成为最有效的应变。初步鉴定分离株S5作为planococcus物种,强调了其在生物修复中的商业应用的潜力。鉴于其效率,进一步的研究应着重于优化环境参数,并扩大工业应用的脱色过程。成功实施这种微生物方法可以提供

自组装刺激激活协调纳米颗粒用于癌症免疫疗法和疫苗应用
支持信息可在https://pubs.acs.org/doi/10.1021/acsnano.3c11374免费获得支持信息。在图S1 – S15中显示的是CDA不同金属离子不同配位行为的照片(图S1); CDA – ZN – MN晶体的EDAX映射(图S2);局部注入CDA或CDA – Zn -MN后,小鼠的血清细胞因子水平(图S3);肿瘤内CDA或CDA – ZN -MN在皮下B16F10黑色素瘤中的治疗作用(图S4); ZMCP的EDAX映射(图S5); PBS中ZMCP,ZCP和MCP的表征(图S6); I.T.后肿瘤中的药物保留注射ZMCP(图S7);肿瘤微环境在肿瘤内注射后对ZMCP调节(图S8);肿瘤内CDA – ZN – MN和ZMCP的治疗作用(图S9);静脉注射后24小时在肿瘤中的药物积累 注射ZMCP(图S10); I.T.的治疗作用 和i.v. 注射ZMCP治疗,并与皮下B16F10黑色素瘤相结合的免疫检查点阻断(图S11); ZMCP-RBD针对RBD序列诱导有效的T细胞反应(图S12);皮下注射后ZMCP的保留和淋巴结排干(图S13); I.T. ZMCP的安全评估 或i.v. 注射(图S14); ZMCP-RBD和ZMCP-S1疫苗接种后主要器官的组织学分析(图S15)(PDF)注射ZMCP(图S7);肿瘤微环境在肿瘤内注射后对ZMCP调节(图S8);肿瘤内CDA – ZN – MN和ZMCP的治疗作用(图S9);静脉注射后24小时在肿瘤中的药物积累注射ZMCP(图S10); I.T.的治疗作用 和i.v. 注射ZMCP治疗,并与皮下B16F10黑色素瘤相结合的免疫检查点阻断(图S11); ZMCP-RBD针对RBD序列诱导有效的T细胞反应(图S12);皮下注射后ZMCP的保留和淋巴结排干(图S13); I.T. ZMCP的安全评估 或i.v. 注射(图S14); ZMCP-RBD和ZMCP-S1疫苗接种后主要器官的组织学分析(图S15)(PDF)注射ZMCP(图S10); I.T.的治疗作用和i.v.注射ZMCP治疗,并与皮下B16F10黑色素瘤相结合的免疫检查点阻断(图S11); ZMCP-RBD针对RBD序列诱导有效的T细胞反应(图S12);皮下注射后ZMCP的保留和淋巴结排干(图S13); I.T. ZMCP的安全评估或i.v.注射(图S14); ZMCP-RBD和ZMCP-S1疫苗接种后主要器官的组织学分析(图S15)(PDF)

目录
内容实验细节图S1。使用0.15m钠( - ) - dibenzoyl-l-tartrate的洗脱完成了L,L-1 4+和D,D,D,D-1 4+的对映体分离的示例。图S2。 使用阳离子 - 交换色谱法分辨出L,L -L -1 4+,D,D,D -1 4+和D,L -1 4+的圆形二色光谱。 表S1。 [D,D -1] Cl 4的晶体数据摘要。 表S2。 [L,L -1] Cl 4的晶体数据摘要。 图S3。 用于[D,D -1] Cl 4晶体结构图S4的阳离子的热椭圆形图。 用于[L,L -1]阳离子的热椭圆形图(PF 6)4晶体结构图S5。 在将DNA逐渐滴定到过量的情况下,涉及L,L -1 4+(5µm)的水缓冲液滴定的示例(25°C时PH7.0)。 顶部:在5mm Tris中添加CT-DNA,25mm NaCl。 底部:在添加人端粒序列时,HTS,(D [Ag 3(T 2 Ag 3)3])在缓冲液中(10 mmKH 2 PO 4 /k 2 HPO 4,1MM k 2 EDTA在50–200 mm kCl中)。 用HTS( - )L,L -1 4+(5µm)的L,L -1 4+(5μm)的最大发光强度。 与CT -DNA( - )的等效滴定在实验误差中对于D,D -1相同。 显示了与HTS( - )的D,D -1(5µM)在等效滴定上获得的最大发射强度的示例。 箭头指示每个发射图S7的L最大值。 用L,L -1 4+(Lambda),D,D,D -1 4+(Delta)和D,L -1 4+(MESO)在MTT分析中获得的细胞活力数据示例。 图S8。 图S9。图S2。使用阳离子 - 交换色谱法分辨出L,L -L -1 4+,D,D,D -1 4+和D,L -1 4+的圆形二色光谱。表S1。 [D,D -1] Cl 4的晶体数据摘要。 表S2。 [L,L -1] Cl 4的晶体数据摘要。 图S3。 用于[D,D -1] Cl 4晶体结构图S4的阳离子的热椭圆形图。 用于[L,L -1]阳离子的热椭圆形图(PF 6)4晶体结构图S5。 在将DNA逐渐滴定到过量的情况下,涉及L,L -1 4+(5µm)的水缓冲液滴定的示例(25°C时PH7.0)。 顶部:在5mm Tris中添加CT-DNA,25mm NaCl。 底部:在添加人端粒序列时,HTS,(D [Ag 3(T 2 Ag 3)3])在缓冲液中(10 mmKH 2 PO 4 /k 2 HPO 4,1MM k 2 EDTA在50–200 mm kCl中)。 用HTS( - )L,L -1 4+(5µm)的L,L -1 4+(5μm)的最大发光强度。 与CT -DNA( - )的等效滴定在实验误差中对于D,D -1相同。 显示了与HTS( - )的D,D -1(5µM)在等效滴定上获得的最大发射强度的示例。 箭头指示每个发射图S7的L最大值。 用L,L -1 4+(Lambda),D,D,D -1 4+(Delta)和D,L -1 4+(MESO)在MTT分析中获得的细胞活力数据示例。 图S8。 图S9。表S1。[D,D -1] Cl 4的晶体数据摘要。表S2。 [L,L -1] Cl 4的晶体数据摘要。 图S3。 用于[D,D -1] Cl 4晶体结构图S4的阳离子的热椭圆形图。 用于[L,L -1]阳离子的热椭圆形图(PF 6)4晶体结构图S5。 在将DNA逐渐滴定到过量的情况下,涉及L,L -1 4+(5µm)的水缓冲液滴定的示例(25°C时PH7.0)。 顶部:在5mm Tris中添加CT-DNA,25mm NaCl。 底部:在添加人端粒序列时,HTS,(D [Ag 3(T 2 Ag 3)3])在缓冲液中(10 mmKH 2 PO 4 /k 2 HPO 4,1MM k 2 EDTA在50–200 mm kCl中)。 用HTS( - )L,L -1 4+(5µm)的L,L -1 4+(5μm)的最大发光强度。 与CT -DNA( - )的等效滴定在实验误差中对于D,D -1相同。 显示了与HTS( - )的D,D -1(5µM)在等效滴定上获得的最大发射强度的示例。 箭头指示每个发射图S7的L最大值。 用L,L -1 4+(Lambda),D,D,D -1 4+(Delta)和D,L -1 4+(MESO)在MTT分析中获得的细胞活力数据示例。 图S8。 图S9。表S2。[L,L -1] Cl 4的晶体数据摘要。 图S3。 用于[D,D -1] Cl 4晶体结构图S4的阳离子的热椭圆形图。 用于[L,L -1]阳离子的热椭圆形图(PF 6)4晶体结构图S5。 在将DNA逐渐滴定到过量的情况下,涉及L,L -1 4+(5µm)的水缓冲液滴定的示例(25°C时PH7.0)。 顶部:在5mm Tris中添加CT-DNA,25mm NaCl。 底部:在添加人端粒序列时,HTS,(D [Ag 3(T 2 Ag 3)3])在缓冲液中(10 mmKH 2 PO 4 /k 2 HPO 4,1MM k 2 EDTA在50–200 mm kCl中)。 用HTS( - )L,L -1 4+(5µm)的L,L -1 4+(5μm)的最大发光强度。 与CT -DNA( - )的等效滴定在实验误差中对于D,D -1相同。 显示了与HTS( - )的D,D -1(5µM)在等效滴定上获得的最大发射强度的示例。 箭头指示每个发射图S7的L最大值。 用L,L -1 4+(Lambda),D,D,D -1 4+(Delta)和D,L -1 4+(MESO)在MTT分析中获得的细胞活力数据示例。 图S8。 图S9。[L,L -1] Cl 4的晶体数据摘要。图S3。用于[D,D -1] Cl 4晶体结构图S4的阳离子的热椭圆形图。用于[L,L -1]阳离子的热椭圆形图(PF 6)4晶体结构图S5。在将DNA逐渐滴定到过量的情况下,涉及L,L -1 4+(5µm)的水缓冲液滴定的示例(25°C时PH7.0)。顶部:在5mm Tris中添加CT-DNA,25mm NaCl。底部:在添加人端粒序列时,HTS,(D [Ag 3(T 2 Ag 3)3])在缓冲液中(10 mmKH 2 PO 4 /k 2 HPO 4,1MM k 2 EDTA在50–200 mm kCl中)。用HTS( - )L,L -1 4+(5µm)的L,L -1 4+(5μm)的最大发光强度。与CT -DNA( - )的等效滴定在实验误差中对于D,D -1相同。显示了与HTS( - )的D,D -1(5µM)在等效滴定上获得的最大发射强度的示例。箭头指示每个发射图S7的L最大值。用L,L -1 4+(Lambda),D,D,D -1 4+(Delta)和D,L -1 4+(MESO)在MTT分析中获得的细胞活力数据示例。图S8。图S9。lambda堆叠实验显示了活的MCF -7细胞中A)D,D -1 4+和L -1 4+的发射曲线。MCF7细胞的CLSM图像使用两个单独的检测通道,分别为670-700 nm(红色)和630-640 nm(黄色),对于D,D,D -1 4+(TOP)和L,L,L -1 4+(底部)。

礁区是整个Phanerokoic
图2:phanerogiac海洋无脊椎动物动物多样性的差异(红色)和非雷夫支持(黑色)区域(黑色)区域(相等的六边形/五角形网格细胞)与所有面板的间隔为1000 km)。排除了明确识别为代表无标准或偏低的存款的收集,也排除了没有有关刻板风格的信息的藏品(见图S7用于使用其他筛选标准的模式)。虚线表示地质时代之间的边界。注意对数Y轴。对于面板A – B和D – F,交叉代表单个网格细胞区域的SQS多样性估计值,而趋势线代表地质时期区域多样性的中值和四分位数。(a)具有空间标准化的phanerogiac海洋动物多样性,对珊瑚礁支持和非雷夫支持区域的对比模式。请注意,在珊瑚礁支持区域中,自奥陶纪以来的多样性水平广为人知,没有长期的世俗趋势证据。从奥陶纪到最新的白垩纪相似,当时多样性相当快地升至通过新生代维持的新的,更高的水平。但是,这种K/PG的增加与腹足类和非污染沉积物密切相关(见图s6)。(b)使用Berger-Parker优势指数(35),在珊瑚礁支持和非Reef支持的网格细胞中估算的均匀度。面板(D – F)显示了Sepkoski进化动物的模式。(c)使用相同的时间箱通过phanerozoic的珊瑚礁支撑和非冰河支撑细胞计数。(d)Cambrian动物群(Trilobita,Linguliformea,Graptolithina,Conodonta); (e)现代动物区系(Anthozoa,ostracoda,Rhynchonelliformea,头孢菌,Crinoidea); (f)现代动物群(Bryozoa,Bivalvia,Gastropoda,Echinoidea,Chondrichthyes)。