XiaoMi-AI文件搜索系统
World File Search SystemSCID

患者来源的非小细胞肺癌异种移植反映了复杂的肿瘤异质性
摘要 目的:患者来源的异种移植(PDX)模型在临床前和转化应用中显示出巨大的潜力,但它们与原发性肿瘤在表型、遗传和药效动力学异质性的一致性尚未得到充分研究。本研究旨在建立非小细胞肺癌(NSCLC)的PDX库,并进一步阐明它是否能保留患者肿瘤内和肿瘤间的异质性。方法:将75例手术切除的NSCLC标本植入免疫缺陷的NOD/SCID小鼠体内。在成功建立NSCLC PDX模型的基础上,我们采用苏木精和伊红染色和免疫组织化学染色比较了癌组织和PDX模型之间波形蛋白、Ki67、EGFR和PD-L1蛋白的表达。此外,我们检测了原发性肿瘤和PDX代之间的全基因表达谱。我们还对 17 个第一代异种移植瘤进行了全外显子组测序 (WES) 分析,以进一步评估 PDX 是否保留了患者的异质性。最后,使用紫杉醇、顺铂、阿霉素、阿替利珠单抗、阿法替尼和 AZD4547 评估 PDX 模型对标准治疗药物的反应。结果:成功开发了大量可连续移植的 NSCLC PDX 模型。PDX 异种移植瘤的组织学和病理免疫组织化学与患者的肿瘤样本一致。WES 和 RNA-seq 进一步证实 PDX 准确复制了原发性肿瘤的分子异质性。与临床患者相似,PDX 模型对标准治疗(包括化疗、靶向和免疫治疗)的反应不同。结论:我们建立的 NSCLC PDX 模型忠实地再现了分子、组织病理学和治疗特征以及相应的肿瘤异质性,为药物筛选、生物标志物发现和转化研究提供了临床相关平台。关键词 患者来源的异种移植 (PDX);非小细胞肺癌 (NSCLC);肿瘤异质性

1 crispr-cas9-pold3融合的快速基因组编辑...
1。Hustedt N,DurocherD。通过细胞周期对DNA修复的控制。自然细胞生物学19,1-9(2017)。2。Miyaoka Y等。对HDR和NHEJ的系统定量揭示了基因组,核酸酶和细胞类型对基因组编辑的影响。科学报告6,23549(2016)。3。Roth TL等。用非病毒基因组靶向重编程人T细胞功能和特异性。自然559,405-409(2018)。4。Yang S,Li S,Li X-J。 缩短CAS9的半衰期具有其基因编辑能力并降低神经元毒性。 细胞报告25,2653-2659。 E2653(2018)。 5。 Haapaniemi E,Botla S,Persson J,Schmierer B,Taipale J. CRISPR – CAS9基因组编辑诱导p53介导的DNA损伤响应。 自然医学24,927-930(2018)。 6。 savic N等。 DNA修复模板与CRISPR-CAS9核酸酶的共价连接增强了同源指导的修复。 Elife 7,E33761(2018)。 7。 Maruyama T,Dougan SK,Truttmann M,Bilate AM,Ingram JR,Ploegh HL。 抑制非同源端连接的抑制会提高CRIS/CAS9介导的精确[TM:插入]基因组编辑的效率。 自然生物技术33,538(2015)。 8。 Robert F,Barbeau M,éthierS,Dostie J,Pelletier J. DNA-PK的药理抑制作用刺激Cas9介导的基因组编辑。 基因组医学7,93(2015)。 9。 自然通讯9,1-9(2018)。 10。 Gu Y等。Yang S,Li S,Li X-J。缩短CAS9的半衰期具有其基因编辑能力并降低神经元毒性。细胞报告25,2653-2659。 E2653(2018)。5。Haapaniemi E,Botla S,Persson J,Schmierer B,Taipale J. CRISPR – CAS9基因组编辑诱导p53介导的DNA损伤响应。自然医学24,927-930(2018)。6。savic N等。DNA修复模板与CRISPR-CAS9核酸酶的共价连接增强了同源指导的修复。Elife 7,E33761(2018)。7。Maruyama T,Dougan SK,Truttmann M,Bilate AM,Ingram JR,Ploegh HL。抑制非同源端连接的抑制会提高CRIS/CAS9介导的精确[TM:插入]基因组编辑的效率。自然生物技术33,538(2015)。8。Robert F,Barbeau M,éthierS,Dostie J,Pelletier J. DNA-PK的药理抑制作用刺激Cas9介导的基因组编辑。基因组医学7,93(2015)。9。自然通讯9,1-9(2018)。10。Gu Y等。Gu Y等。Riesenberg S,Maricic T.用小分子靶向修复途径会增加多能干细胞中精确的基因组编辑。ku70缺陷小鼠的生长迟缓和漏水的SCID表型。免疫7,653-665(1997)。

同种异体加工的胸腺组织-AGDC(Rethymic®)
简介Rethymic®由培养的胸腺组织(CTT)组成,该组织被处理以保留胸腺上皮细胞并耗尽大多数供体胸腺细胞,然后再将胸膜细胞植入接受者Quadriceps肌肉中的口袋。ctt的作用与正常胸腺组织相似,产生免疫功能的幼稚T细胞(CD3+ CD4+ CD45RA+ CD62L+和CD3+ CD8+ CD8+ CD45RA+ CD62L+细胞),可以迁移到外围,并以CD3+ CD4+ CD4+ CD3+ CD3+ CD8+ CD8+ CD8+细胞迁移。在CTT移植后,受体骨髓干细胞转到移植组织以发展为受体T细胞。受体树突状细胞删除了与树突状细胞过于结合的任何发育中的胸腺细胞或CTT中供体胸上皮的结合。没有T细胞会攻击受体或CTT离开胸腺。 天真的T细胞出现在植入后6个月和12个月之间的外周血中出现(Markert,2022)。 再生剂量取决于组织切片的总表面积和受体的身体表面积(BSA)。 将切片定义为单个滤膜上的内容。 建议的剂量范围是每m 2受体BSA的再生表面积2至22,000 mm 2。 根据制造商预先计算的特定患者(FDA,Rethymic全处方信息,2021年),在单剂量单位中最多提供42片切片。 Markert及其同事(2022)在1993年至2020年中证明了10个前瞻性,单中心的开放标签研究的功效。 总共招募了105名患者并接受了CTT移植。 呼吸衰竭是负责的原因没有T细胞会攻击受体或CTT离开胸腺。天真的T细胞出现在植入后6个月和12个月之间的外周血中出现(Markert,2022)。再生剂量取决于组织切片的总表面积和受体的身体表面积(BSA)。将切片定义为单个滤膜上的内容。建议的剂量范围是每m 2受体BSA的再生表面积2至22,000 mm 2。根据制造商预先计算的特定患者(FDA,Rethymic全处方信息,2021年),在单剂量单位中最多提供42片切片。Markert及其同事(2022)在1993年至2020年中证明了10个前瞻性,单中心的开放标签研究的功效。总共招募了105名患者并接受了CTT移植。呼吸衰竭是负责,除先天性小没有小没有小没有障碍和/或接受了先前的治疗外,有10名患者有诊断。中包括95例。For inclusion, patients had to have athymia as defined by flow cytometry demonstrating a circulating CD3+ CD45RA+CD62L+ T cell count lower than 50/mm 3 or less than 5% of the total T cell count on 2 separate flow cytometry analyses (1 performed within 3 months and 1 performed within 1 month before administration of CTT), unless they were enrolled in the expanded access protocol, according to which the naïve T cell计数可能高于50mm 3。其他关键资格标准包括CDGA或FOXN1缺乏症以及与SCID相关的遗传缺陷。排除标准包括在给药前4周内进行心脏手术,外科医生或麻醉医生确定的手术不良,HIV感染,事先尝试进行免疫结构,呼吸机依赖性和巨细胞病毒(CMV)感染,需要用于需要免疫抑制的患者。免疫抑制是根据对植物凝集素(PHA)的增生反应,无论典型或非典型表型。在疗效分析集(EAS)中,有95例先天性胸肌患者中有93例CDGA诊断和2例FOXN1缺乏症。Kaplan-Meier估计在收到CTT后1年和2年内EA的生存率分别为77%和76%。 EA的中位随访时间为7。6年,范围从0到25。5年。 CTT移植后约1-2年,T细胞计数达到了峰值。Kaplan-Meier估计在收到CTT后1年和2年内EA的生存率分别为77%和76%。EA的中位随访时间为7。6年,范围从0到25。5年。CTT移植后约1-2年,T细胞计数达到了峰值。对于收到CTT后1年活着的患者,中位随访时间为10.9岁的估计存活率为93%。幼稚的T细胞数量从所有患者开始为0,并在2年内增加到最高数量。通过对有丝分裂原和抗原的增生反应研究 T细胞功能。 在大多数患者中, b细胞和NK细胞计数保持在正常范围内,几乎所有患者都能够阻止IgG替代疗法(Markert,2022)。 有105例患者,有32例至少有1例严重不良事件的患者,35例至少有1例威胁生命的不良事件,以及26例与事件有关的死亡。 总共有53例患者被归类为严重,13例被归类为威胁生命,11例患有致命感染。 在CTT移植后的第一年,总共报道了78种新的病毒感染。 105例患者中共有28例死亡;由于CTT移植后进行了SCID诊断,因此认为26例与不良事件有关,另有2例报告在研究后报告。 在移植后的第一年,在28例死亡中,22例死亡(包括13例与感染有关的死亡中的12例)发生,而患者仍然是免疫缺陷的。T细胞功能。b细胞和NK细胞计数保持在正常范围内,几乎所有患者都能够阻止IgG替代疗法(Markert,2022)。有105例患者,有32例至少有1例严重不良事件的患者,35例至少有1例威胁生命的不良事件,以及26例与事件有关的死亡。总共有53例患者被归类为严重,13例被归类为威胁生命,11例患有致命感染。在CTT移植后的第一年,总共报道了78种新的病毒感染。105例患者中共有28例死亡;由于CTT移植后进行了SCID诊断,因此认为26例与不良事件有关,另有2例报告在研究后报告。在移植后的第一年,在28例死亡中,22例死亡(包括13例与感染有关的死亡中的12例)发生,而患者仍然是免疫缺陷的。

使用慢病毒载体的造血干细胞基因治疗的现状...
2021; 26(10):38-43。3)sasaki sumimi inoue takao。 COVID -19疫苗 - 适度的扩散。学术趋势2021; 26(10):31-7。4)Hacein-Bey-Abina S,Pai Sy,Gaspar HB等。用于X连锁严重合并免疫缺陷的修饰γ-逆转录病毒载体。n Engl J Med 2014; 371:1407-17。5)Onodera Masafumi。通过基因组编辑进行基因治疗的进一步发展。日本造血细胞移植协会杂志2018; 7(2):32-9。6)BöckD,Rothgangl T,Villiger L等。在小鼠中的体内质量编辑。 SCI Transl Med 2022; 14:EABL9238。 7)Demeulemeester J,de Rijck J,Gijsbers R,Debyser Z.逆转录病毒Inte-Crimination:地点事项:逆转录病毒Inte磨牙部位选择的机制和后果。 生物评估2015; 37:1202-14。 8)Liang Q,Vlaar EC,Catalano F等。 慢病毒基因治疗可防止鼠绒性疾病中的抗人类酸α-葡萄糖苷酶抗体形成。 mol ther方法Clin Dev 2022; 25:520-32。 9)Cavazzana-Calvo M,Hacein-Bey S,De Saint Basile G等。 人类严重合并免疫缺陷(SCID)-X1疾病的基因疗法。 Science 2000; 288:669-72。 10)Hacein-Bey-Abina S,Le Deist F,Carlier F等。 通过体内基因治疗对X连锁严重的免疫缺陷进行持续校正。 n Engl J Med 2002; 346:1185-93。 11)Howe SJ,Mansour MR,Schwarzwaelder K等。 插入诱变与获得的体细胞突变相结合导致SCID-X1患者基因治疗后的白血病发生。 J Clin Invest 2008; 118:3143-50。 12)Cartier N,Hacein-Bey-Abina S,Bartholomae CC等。在小鼠中的体内质量编辑。SCI Transl Med 2022; 14:EABL9238。7)Demeulemeester J,de Rijck J,Gijsbers R,Debyser Z.逆转录病毒Inte-Crimination:地点事项:逆转录病毒Inte磨牙部位选择的机制和后果。生物评估2015; 37:1202-14。8)Liang Q,Vlaar EC,Catalano F等。慢病毒基因治疗可防止鼠绒性疾病中的抗人类酸α-葡萄糖苷酶抗体形成。mol ther方法Clin Dev 2022; 25:520-32。9)Cavazzana-Calvo M,Hacein-Bey S,De Saint Basile G等。人类严重合并免疫缺陷(SCID)-X1疾病的基因疗法。Science 2000; 288:669-72。 10)Hacein-Bey-Abina S,Le Deist F,Carlier F等。 通过体内基因治疗对X连锁严重的免疫缺陷进行持续校正。 n Engl J Med 2002; 346:1185-93。 11)Howe SJ,Mansour MR,Schwarzwaelder K等。 插入诱变与获得的体细胞突变相结合导致SCID-X1患者基因治疗后的白血病发生。 J Clin Invest 2008; 118:3143-50。 12)Cartier N,Hacein-Bey-Abina S,Bartholomae CC等。Science 2000; 288:669-72。10)Hacein-Bey-Abina S,Le Deist F,Carlier F等。通过体内基因治疗对X连锁严重的免疫缺陷进行持续校正。n Engl J Med 2002; 346:1185-93。11)Howe SJ,Mansour MR,Schwarzwaelder K等。插入诱变与获得的体细胞突变相结合导致SCID-X1患者基因治疗后的白血病发生。J Clin Invest 2008; 118:3143-50。12)Cartier N,Hacein-Bey-Abina S,Bartholomae CC等。造血细胞基因疗法在X连锁性肾上腺肌营养不良症中使用慢病毒载体。Science 2009; 326:818-23。 13)Biffi A,Montini E,Lorioli L等。 慢病毒造血干细胞Science 2009; 326:818-23。13)Biffi A,Montini E,Lorioli L等。慢病毒造血干细胞
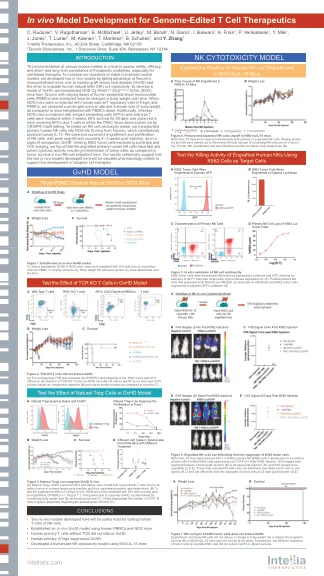
基因组编辑 T 细胞治疗的体内模型开发
实施各种小鼠模型对于评估治疗方式的安全性、有效性以及短期和长期持久性至关重要,尤其是对于基于细胞的疗法。为了增加我们可行的人源化小鼠模型库,我们利用 Taconic 的免疫缺陷小鼠开发了两种体内模型:一种用于监测移植物抗宿主病 (GvHD),另一种用于评估人类自然杀伤 (NK) 细胞的细胞毒性。为了开发 GvHD 模型,我们向 Taconic 的 NOD.Cg- Prkdc scid Il2rg tm1Sug / JicTac (NOG) 小鼠移植了不同剂量的人类外周血单核细胞 (PBMC),并监测小鼠体重随时间的变化。当 NOG 小鼠同时注射人类天然 T 调节细胞 (nTregs) 和 PBMC 时,与仅移植 PBMC 的小鼠相比,我们观察到存活率延长且体重减轻较慢。此外,同时注射抗原呈递细胞 (APC) 和野生型 T 细胞的 NOG 小鼠在 3 周内就奄奄一息,而接受 APC 加 T 细胞(其中 TRAC 基因座通过 CRISPR / Cas9 编辑被敲除)的小鼠在 90 天内存活率达到 80%。为了建立 NK 细胞毒性模型,我们将原代人类 NK 细胞移植到 Taconic 的 NOG-hIL15 小鼠中,该小鼠组成性产生人类 IL-15。我们观察到 NK 细胞的成功植入和增殖,峰值植入发生在注射后 4 - 5 周,没有异种 GvHD 的迹象。利用表达荧光素酶的 K562 肿瘤细胞和 IVIS 成像,我们发现植入的原代人类 NK 细胞具有快速而强大的细胞毒活性,与非 NK 细胞植入小鼠的肿瘤存活率相比,可消除肿瘤细胞。我们的研究结果共同表明,这里开发的两种体内模型将成为支持过继细胞疗法发展的有价值的药理学模型。
一周的课程粘膜皮肤病
1 BBC在线新闻。基因疗法保存的“泡泡男孩”。2002年4月3日,http://news.bbc.co.uk/1/hi/health/19069999.stm(2004年8月20日访问)。2 Cavazzana-Calvo M,Hacein-Bey S,De Saint Basile G,Gross F,Yvon E,Nusbaum P等。人类严重联合免疫耐智能(SCID)-X1疾病的基因疗法。Science 2000; 288:669-72:3 Aiuti A,Slavin S,Aker M,Ficara F,Deola S,Mortellaro A等。通过干细胞基因疗法对ADA-SCID进行纠正,并结合非甲莫氏菌度调节。Science 2002; 296:2410-3 .. 4 Hacein-Bey-Abina S,Von Kalle C,Schmidt M,McCormack MP,Wulffraat N,Leboulch P等。SCID-X1基因治疗后两名患者的LMO2相关克隆T细胞增殖。Science 2003; 302:415-9。 5 Kay MA,Manno CS,Ragni MV,Larson PJ,Couto LB,McClelland A等。 证据证明了用AAV载体治疗的血友病患者中IX因子IX的表达。 自然基因2000; 24:257-61。 6 Lehrman S.基因治疗死亡后提出了质疑的病毒治疗。 自然1999; 401517-8。 7 Kay MA,Nakai H.研究AAV矢量的安全。 自然2003; 424:251。 8 Bushman F.靶向逆转录病毒整合。 mol ther 2000; 6。 9 Olivares EC,Hollis RP,Chalberg TW,Meuse L,Kay MA,Calos MP。 位点特异性基因组整合产生小鼠的治疗因子IX水平。 自然生物技术2002; 20:1124-8。 10 Ries,S和Kirn,W M. Onyx-015:复制选择性腺病毒的作用机理和临床潜力。 br J Cancer 2002; 86:5-11。Science 2003; 302:415-9。5 Kay MA,Manno CS,Ragni MV,Larson PJ,Couto LB,McClelland A等。 证据证明了用AAV载体治疗的血友病患者中IX因子IX的表达。 自然基因2000; 24:257-61。 6 Lehrman S.基因治疗死亡后提出了质疑的病毒治疗。 自然1999; 401517-8。 7 Kay MA,Nakai H.研究AAV矢量的安全。 自然2003; 424:251。 8 Bushman F.靶向逆转录病毒整合。 mol ther 2000; 6。 9 Olivares EC,Hollis RP,Chalberg TW,Meuse L,Kay MA,Calos MP。 位点特异性基因组整合产生小鼠的治疗因子IX水平。 自然生物技术2002; 20:1124-8。 10 Ries,S和Kirn,W M. Onyx-015:复制选择性腺病毒的作用机理和临床潜力。 br J Cancer 2002; 86:5-11。5 Kay MA,Manno CS,Ragni MV,Larson PJ,Couto LB,McClelland A等。证据证明了用AAV载体治疗的血友病患者中IX因子IX的表达。自然基因2000; 24:257-61。6 Lehrman S.基因治疗死亡后提出了质疑的病毒治疗。自然1999; 401517-8。7 Kay MA,Nakai H.研究AAV矢量的安全。 自然2003; 424:251。 8 Bushman F.靶向逆转录病毒整合。 mol ther 2000; 6。 9 Olivares EC,Hollis RP,Chalberg TW,Meuse L,Kay MA,Calos MP。 位点特异性基因组整合产生小鼠的治疗因子IX水平。 自然生物技术2002; 20:1124-8。 10 Ries,S和Kirn,W M. Onyx-015:复制选择性腺病毒的作用机理和临床潜力。 br J Cancer 2002; 86:5-11。7 Kay MA,Nakai H.研究AAV矢量的安全。自然2003; 424:251。8 Bushman F.靶向逆转录病毒整合。mol ther 2000; 6。9 Olivares EC,Hollis RP,Chalberg TW,Meuse L,Kay MA,Calos MP。位点特异性基因组整合产生小鼠的治疗因子IX水平。自然生物技术2002; 20:1124-8。10 Ries,S和Kirn,W M. Onyx-015:复制选择性腺病毒的作用机理和临床潜力。br J Cancer 2002; 86:5-11。11 Cavazzan-Calvo M,Thrusher A,Mavilio F.自然2004; 427:779-81。

人性化小鼠中人类癌异种移植物的表征
针对人类免疫系统的药物的抽象背景临床前评估对肿瘤学研究人员提出了挑战。由于人性化小鼠的商业引入,抗肿瘤功效和药效研究现在可以用带有人类免疫系统成分的小鼠中的人类癌细胞进行。但是,这些模型的开发和表征对于了解哪种模型最适合不同的代理是必要的。我们表征了A375,A549,CAKI-1,H1299,H1975,HCC827,HCC827,HCT116,KU-19-19,MDA-MB-231,MDA-MB-231和RKO Human Cell Cell Cell Senografts CD34 + CD34 +人类糖尿病型糖尿病生长量的Tumore Lagrig ravention tumune tumune tumune tumune tumune tumune tumune tumune 1 (PD-L1)表达和对抗PD-L1治疗的反应。使用流式细胞术和免疫组织化学进行免疫细胞分析。使用atezolizumab进行了人源化异种移植模型对PD-L1治疗的抗肿瘤反应。结果我们发现,与其他模型相比,与模型中的脾脏和肿瘤中的CD4 +和CD8 + T细胞组成不同,A375,CAKI-1,MDA-MB-231和HCC827含有较高的肿瘤内频率,与其他模型相比,CD4 +和CD8 + T细胞含有较高的肿瘤内频率。我们证明,每个模型内的免疫细胞浸润水平都受肿瘤而不是干细胞供体的强烈影响。许多肿瘤模型表现出丰富的髓样细胞,B细胞和树突状细胞。RKO和MDA-MB-231肿瘤包含PD-L1 +肿瘤细胞的最高表达。RKO和MDA-MB-231肿瘤包含PD-L1 +肿瘤细胞的最高表达。模型对atezolizumab的抗肿瘤反应与CD4 +和CD8 +肿瘤浸润淋巴细胞(TILS)的水平呈正相关。结论这些数据表明,肿瘤内部因素会影响肿瘤和脾脏内的免疫细胞库,并且TIL频率是确定人道化小鼠中肿瘤异种移植物中对抗PD-L1反应的关键因素。这些数据还可能有助于选择肿瘤模型,以测试新型免疫肿瘤或肿瘤定向剂的抗肿瘤活性。

2024年东南化学生物学和药物发现...
迫切需要具有增强效力和特异性的新型抗病毒药物来治疗水痘带状疱疹病毒 (VZV) 和单纯疱疹病毒 (1 和 2) 感染。L-BHDU(β-L-1-[5-(E-2-溴乙烯基)-2-(羟甲基)-1,3-(二氧杂环戊烷-4-基)]尿嘧啶)在培养细胞和人源化 SCID 小鼠中对水痘带状疱疹病毒 (VZV) 具有高度活性。此外,L-BHDU 对 HSV1 也具有高度活性,对 HSV2 具有中等活性(EC 50 7 µM)。我们采用前药方法合成了带有双(新戊酰氧甲基)基团的单磷酸盐前药 POM-L-BHDU-MP,以提高其药理特性,同时保留抗病毒活性(VZV EC 50 0.04 µM;HSV1 EC 50 0.03 µM;CC 50 >100 µM)。在体内实验中,我们在皮肤器官培养物和小鼠中评估了 POM-L-BHDU-MP 对抗 VZV 和 HSV1 的作用,并研究了其药代动力学和分布特性。可可脂中的 POM-L-BHDU(0.2% 顶部)可防止 VZV 或 HSV1 扩散,并且对人体皮肤外植体无毒。在 NuSkin 小鼠模型中,POM-L-BHDU-MP 可减少经皮下和口服途径的 VZV 扩散(45、22.4、11.3 mg/kg)并且耐受性良好。在 BALB/c 小鼠皮肤侧腹模型中,POM-L-BHDU-MP(22.4 mg/kg 口服)减轻了 HSV1 引起的体重减轻,更多研究正在进行中。给小鼠口服或静脉注射 POM-L-BHDU-MP,然后用 LC-MS/MS 分析其血浆和器官。POM-L-BHDU-MP 迅速转化为 L-BHDU,口服生物利用度高。血浆中的 L-BHDU(22.5 mg/kg 口服)达到 C max 为 10 ± 2.5 µg/mL,T max 为 0.85 小时;半衰期为 5-6 小时。L-BHDU 分布在小鼠器官中,包括大脑和脑脊液。 L-BHDU 的磷酸化代谢物在未感染的小鼠中是痕量的,而 L-BHDU 的二磷酸盐和三磷酸盐形式在 VZV 感染的人类皮肤异种移植中为 30-50 µM,超过了 EC 90 。总体而言,POM-L-BHDU-MP 是 L-BHDU 的强效前药,是一种有前途的核苷酸类似物,可用于治疗 VZV 和 HSV1 感染。

2024东南化学生物学和药物发现...
迫切需要具有增强效力和特异性的新抗病毒药物来治疗水痘带状疱疹病毒(VZV)和单纯疱疹病毒(1&2)感染。l-bhdu(β-L-1- [5-(E-2-溴叶烯基)-2-(羟甲基)-1,3-(二恶英-4-基)] uracil在培养的细胞和人型scid小鼠中对Vericella Zoster病毒(VZV)具有高度活性。此外,L-BHDU对HSV1也具有很高的活性,并且对HSV2(EC 50 7 µM)表现出了中等活性。使用前药方法,我们将单磷酸前药POM-L-BHDU-MP与BIS(Pivaloyloxymethyl)组合成,以改善其药理性能,同时保留抗病毒活性(VZV EC 50 0.04 µM; HSV1 EC 50 0.03 µm; HSV1 EC 50 0.03 µm; CC 50> 100> 100> 100> 100> 100> 100 µm)。在体内,对皮肤器官培养物和小鼠的VZV和HSV1评估了POM-L-BHDU-MP,并研究了药代动力学和分布性能。pom-l-bhdu(顶部为0.2%)可以防止VZV或HSV1扩散,并且对人皮外植体无毒。在Nuskin小鼠模型中,POM-L-BHDU-MP通过皮下和口服路线(45、22.4、11.3 mg/kg)降低了VZV扩散,并且耐受性良好。在BALB/C小鼠皮肤侧面模型中,POM-L-BHDU-MP(22.4 mg/kg PO)减少了HSV1诱导的体重减轻,并且正在进行更多的研究。小鼠口服或静脉注射POM-L-BHDU-MP,并通过LC-MS/MS分析其血浆和器官。POM-L-BHDU-MP迅速转化为L-BHDU,口服生物利用度很高。l-BHDU(22.5 mg/kg PO)达到10±2.5 µg/ml的C最大为0.85 h;半衰期是5-6小时。 L-BHDU分布在小鼠器官中,包括大脑和脑脊液。L-BHDU的磷酸化代谢产物是未感染的小鼠的痕迹,而L-BHDU的Di-和三磷酸形式为30-50 µm,在VZV感染的人类皮肤异种移植物中,超过EC 90。总体而言,POM-L-BHDU-MP是L-BHDU的有效前药,这是一种可用于治疗VZV和HSV1感染的有前途的核苷酸类似物。

自适应免疫和自身免疫|摘要
001 Adaptive Immunity and Autoimmunity Human scalp hair follicles are protected from alopecia areata in vivo by regulatory γδ T cells A Keren, 1 N Goldstein, 1 M Bertolini, 2 R Kassem, 3 R Paus 4,5 and A Gilhar 1 1 Technion – Israel Institute of Technology, Haifa, Israel, 2 Monasterium Laboratory, a Qima Life Sciences Company,Münster,德国,3个皮肤科系,Sheba医学中心,特拉维夫大学,特拉维夫,以色列4,迈阿密大学皮肤病学系4,佛罗里达州米亚,佛罗里达大学和5次沙皇 - 皮肤和头发创新,德国汉堡,德国,德国Foxp3+调节性T细胞(Tregs)cr anterant的角色扮演不受控制的角色。与AREATA(AA)受影响的头皮相比,健康头皮的毛囊(HFS)中存在毛囊(HFS)的数量明显更高,但它们在AA发病机理中的作用尚不清楚。为了研究其在体内AA发育中的作用,我们使用了AA人源化的小鼠模型,其中健康的人体皮肤被异种移植到SCID/米色小鼠上,并注射CD8+/ NKG2D+TCELLS来诱导AA样病变。γδTreg是由用IL-2/IL-15/唑来膦酸盐/TGF-beta培养的PBMC在体外产生的。预防性注射γδTregs侵害了免于AA病变形成的异种移植物,而注射到现有病变中则促进了头发再生,减少了叶面浸润并恢复了HF免疫特权(IP)。与“压力”(与CD8+/NKG2D+TCELLS共同培养)的γδTreg孵育人类头皮Hfs ex Vivo导致预防过早脱发,头发矩阵矩阵增殖增殖增加,并减少了IP的collapeians(Mica MORPAUSSIAN)(MICA)(MICA)(MICA)(MICA)(MICA)(MICA)(MICA)(MICA)) )。为进一步阐明了AA中γδTreg的治疗潜力的机制,我们注入了IL-10或TGFβ1中和抗体或IgG对照中中和γδTregs与aA病变中的AA疾病,并发现了AA的AA效果,并发现了AA+ cD的AA+γδ+γδ+γδTregs+ Quds+γδ+γ+细胞数和HLA-A,-b,-c和-dr的表达增加,并伴随着αMSH和TGF-β的降低。这项研究强调γδTreg是针对AA病变形成的关键保护性免疫细胞,该细胞主要由IL-10和TGF-β1介导,强调了γδTregs作为基于基于细胞的新型细胞治疗的潜力。