XiaoMi-AI文件搜索系统
World File Search SystemSSDNA

ssDNA/RNA清洁和集中器
产品描述SSDNA/RNA Clean&Eccentator™套件提供了10分钟,可靠的方法,可快速分离,清理和浓度,最多可从双链种类的单个链DNA和/或RNA(例如,基因组DNA)的单个链DNA和/或RNA。此简单过程基于使用唯一的单支无用系统和Zymo-Spin™列技术。单链DNA或RNA(17至200个核苷酸;例如,短转录本,探针,引物)可以使用此试剂盒安全处理和回收。该结果高度浓缩(≥6µL),纯化的DNA/RNA,适用于随后的分子方法,包括PCR,RT/PCR,杂交等。
有效生产和应用ssDNA在CRISPR/ ... div>
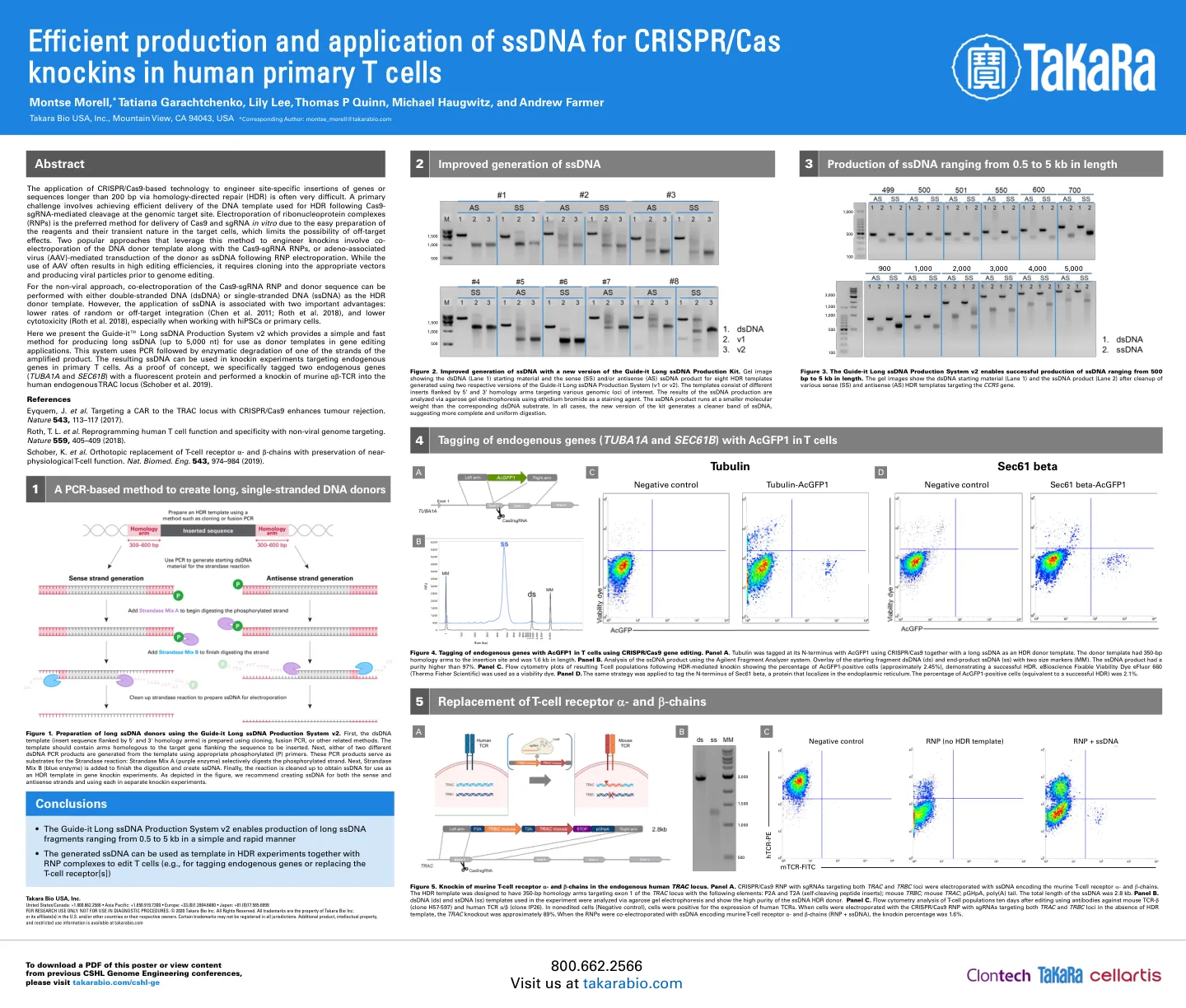
图4。使用CRISPR/CAS9基因编辑在T细胞中用ACGFP1的内源基因标记。面板a。微管蛋白使用CRISPR/CAS9和长ssDNA作为HDR供体模板在其N末端使用ACGFP1标记。供体模板在插入位点具有350 bp的同源性臂,长度为1.6 kb。面板B.使用安捷伦碎片分析仪系统对ssDNA产物进行分析。起始片段dsDNA(ds)和末端产物ssDNA(SS)的覆盖层具有两个大小标记(mm)。ssDNA产品的纯度高于97%。面板C. HDR介导的敲击蛋白后,所得的T细胞群体的流式细胞仪图显示了ACGFP1阳性细胞的百分比(约2.45%),证明了成功的HDR。Ebioscience可固定的生存能力染料Efluor 660(Thermo Fisher Scientific)用作活力染料。面板D.使用相同的策略来标记Sec61β的N末端,Sec61β的N端是一种定位在内质网中的蛋白质。ACGFP1阳性细胞的百分比(相当于成功的HDR)为2.1%。


guide-it™长SSDNA生产系统V2协议-A ...
I.使用单链DNA(SSDNA)而不是双链DNA(DSDNA)作为CRISPR/CAS9敲击实验中同源指导修复(HDR)的供体模板的引言具有几个重要优势。ssDNA在传递到靶细胞时不会触发强烈的细胞毒性反应,与DSDNA不同,将随机整合到基因组中的可能性要小得多(Roth等人,2018年)。对于涉及较长SSDNA的应用,例如用荧光记者标记内源基因,通常以具有成本效益的方式生产无错误的长ssDNA链(超过200个基础)是一个挑战。指南长ssDNA生产系统V2(Cat。编号632666)旨在在涉及CRISPR/CAS9或其他基因编辑工具的敲击实验中生产长ssDNA寡素(从500 nt至5,000 nt)作为供体模板。

新型抗危机辅助CRISPR生物传感器,用于独家检测单链DNA(ssDNA)
摘要:核酸分析在疾病诊断和治疗中起重要作用。CRISPR技术的发现为检测核酸的检测提供了新颖而多功能的方法。但是,使用最广泛的CRISPR-CAS12A检测平台缺乏将单链DNA(ssDNA)与双链DNA(DSDNA)区分开的能力。为了克服这一局限性,我们首先采用了抗Crispr蛋白(ACRVA1)来开发一种新型的CRISPR生物传感器,以专门检测ssDNA。在这种传感策略中,ACRVA1切割CRISPR指南RNA(CRRNA)抑制CRISPR-CAS12A系统的裂解活性。只有ssDNA具有募集裂解的crRNA片段以恢复CRISPR-CAS12生物传感器的检测能力,但DSDNA无法实现这一目标。通过测量CRISPR-CAS12A生物传感器的回收裂解活性,我们开发的ACRVA1辅助CRISPR生物传感器能够将ssDNA与dsDNA区分开,为检测SSDNA的检测提供了一种简单可靠的方法。此外,我们证明了我们开发的ACRVA1辅助CRISPR生物传感器,以监测解旋酶的酶促活性并筛选其抑制剂。关键字:基于CRISPR的生物传感器,CAS12A(CPF1)核酸酶,抗Crispr蛋白,ACRVA1,单链DNA(SSDNA)

ssDNA 纳米管用于选择性靶向胶质母细胞瘤并输送阿霉素以提高生存率
有效治疗胶质母细胞瘤仍然是一项艰巨的挑战。治疗药物开发的主要障碍之一是它们无法穿过血脑肿瘤屏障 (BBTB)。局部给药是一种替代方法,但在缺乏靶标选择性的情况下仍可能产生毒性。在这里,我们展示了由 ssDNA 两亲分子自组装形成的纳米管在血清和核酸酶中是稳定的。双侧脑注射后,纳米管在肿瘤中比在正常脑中更易保留,并通过清道夫受体结合和巨胞饮作用被胶质母细胞瘤细胞吸收。静脉注射后,它们穿过 BBTB 并内化到胶质母细胞瘤细胞中。在微小残留疾病模型中,局部给药阿霉素在脾脏和肝脏中显示出毒性迹象。相反,通过纳米管输送阿霉素不会引起全身毒性,并提高小鼠的存活率。我们的结果表明,ssDNA 纳米管是一种很有前途的胶质母细胞瘤药物输送载体。

通过热诱导 ssDNA 重组技术对假单胞菌进行高效多位点基因组编辑
单链 DNA 重组工程在肠道细菌以外物种的基因组编辑中的应用受到重组酶效率和内源性错配修复 (MMR) 系统作用的限制。在这项工作中,我们建立了一个遗传系统,用于在生物技术相关菌株 EM42 的染色体中输入多种变化。为此,设计了 PL / c I857 系统的控制下,rec2 重组酶和 P. putida 的等位基因 mutL E36K PP 的高水平热诱导共转录。短时间热转移循环,然后用一套诱变寡核苷酸进行转化,可产生不同类型的基因组变化,每次修饰的频率高达 10%。相同的方法有助于超级多样化短染色体部分,以创建功能性基因组片段文库——例如核糖体结合位点。这些结果使得假单胞菌基因组工程的多重化成为可能,这是这种重要合成生物学底盘代谢重编程所必需的。

使用长ssDNA模板进行同源指导修复的IPS细胞中有效的基因敲击蛋白
CRISPR/CAS技术的常见应用涉及工程基因敲击素,其中DNA序列被取代或插入特定的基因组基因座。In contrast with CRISPR-mediated indels, which result from the error-prone non-homologous end joining (NHEJ) pathway, gene knockins are often engineered via homology-directed repair (HDR), typically through the use of CRISPR reagents (Cas enzyme and guide RNA) in tandem with a DNA template that shares homology with the target site and encodes for the desired modification (Hsu et al., 2014;图1,下面)。用于HDR的模板可以是双链DNA(DSDNA,线性或质粒)或单链DNA(SSDNA),并且最近的发现表明,修复机制取决于使用的模板类型而变化。 dsDNA触发了一种反映减数分裂同源重组(HR)的RAD51依赖性机制,而HDR涉及ssDNA(称为单链模板修复或SSTR)是Rad51独立的,并且需要多个组件,并且需要多个组成部分的Fanconi Anemia Anemia(FA)维修路径(RICHARDARDSON ERATHEWAY(RICHARDARSEN)等。
数字核酸记忆:新型P11453.1 DNA折纸的脚手架的缩放ssDNA合成
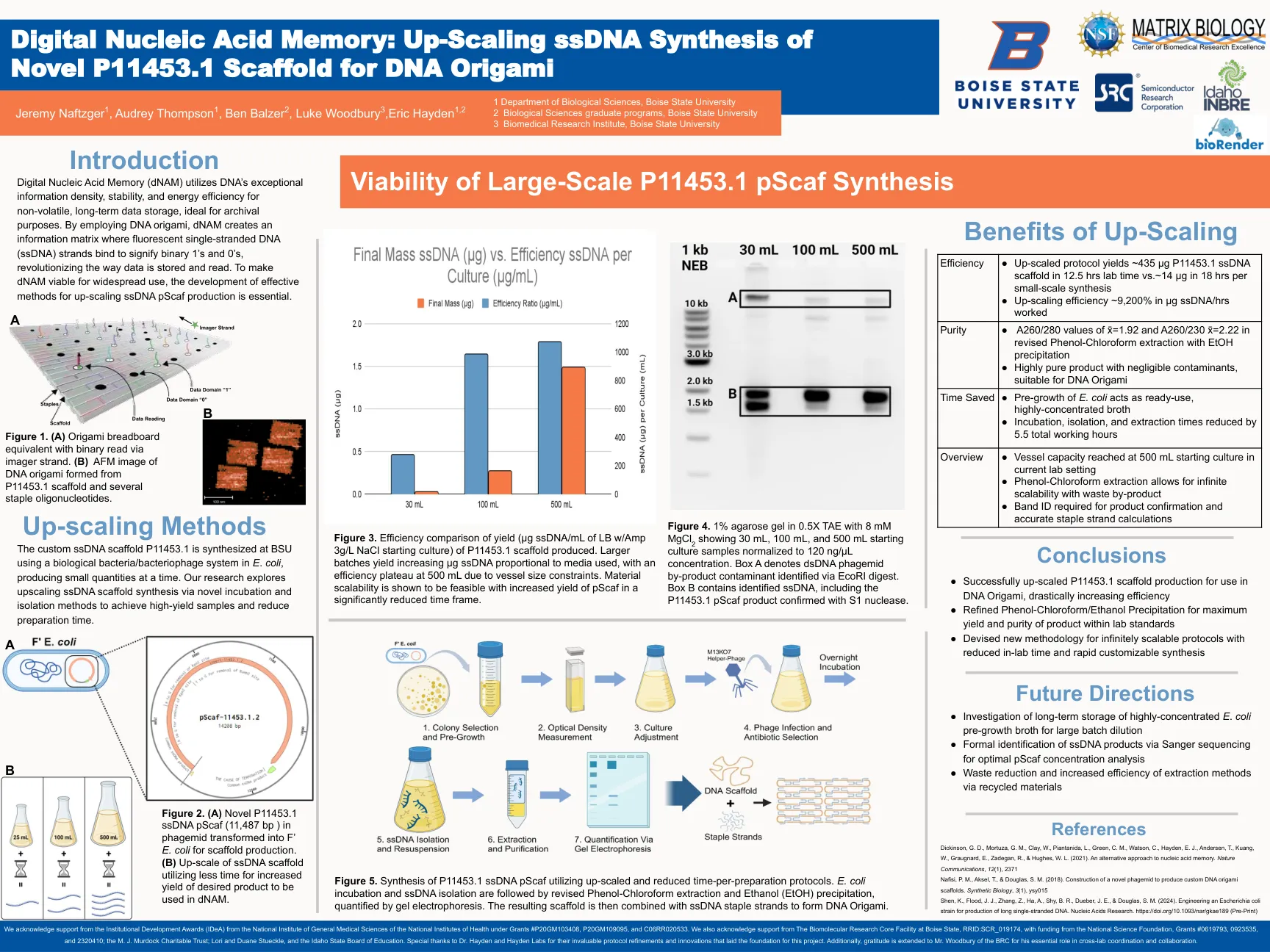
数字核酸记忆(DNAM)利用DNA的非挥发性,长期数据存储的DNA的特殊信息密度,稳定性和能源效率,非常适合档案目的。通过使用DNA折纸,DNAN创建了一个信息矩阵,其中荧光单链DNA(SSDNA)链结合以表示二进制1和0,从而革新了数据存储和读取的方式。使DNAM适用于广泛使用,开发了提高SSDNA PSCAF生产的有效方法是必不可少的。

将 dsDNA 与 ssDNA Oligo 和 NEBuilder HiFi DNA 组装连接起来,创建 sgRNA-Cas9 表达载体
摘要 本应用说明展示了将 sgRNA 序列插入 9.5 kb 载体进行靶向 DNA 组装的便利性。与必须合成并重新退火两个寡核苷酸的传统克隆方法不同,此新方案提供了一种简单的方法来设计寡核苷酸并将其与所需载体组装。NEBuilder HiFi DNA 组装主混合物比传统方法有了显著的改进,特别是在节省时间、易于使用和成本方面。