XiaoMi-AI文件搜索系统
World File Search SystemSTK11
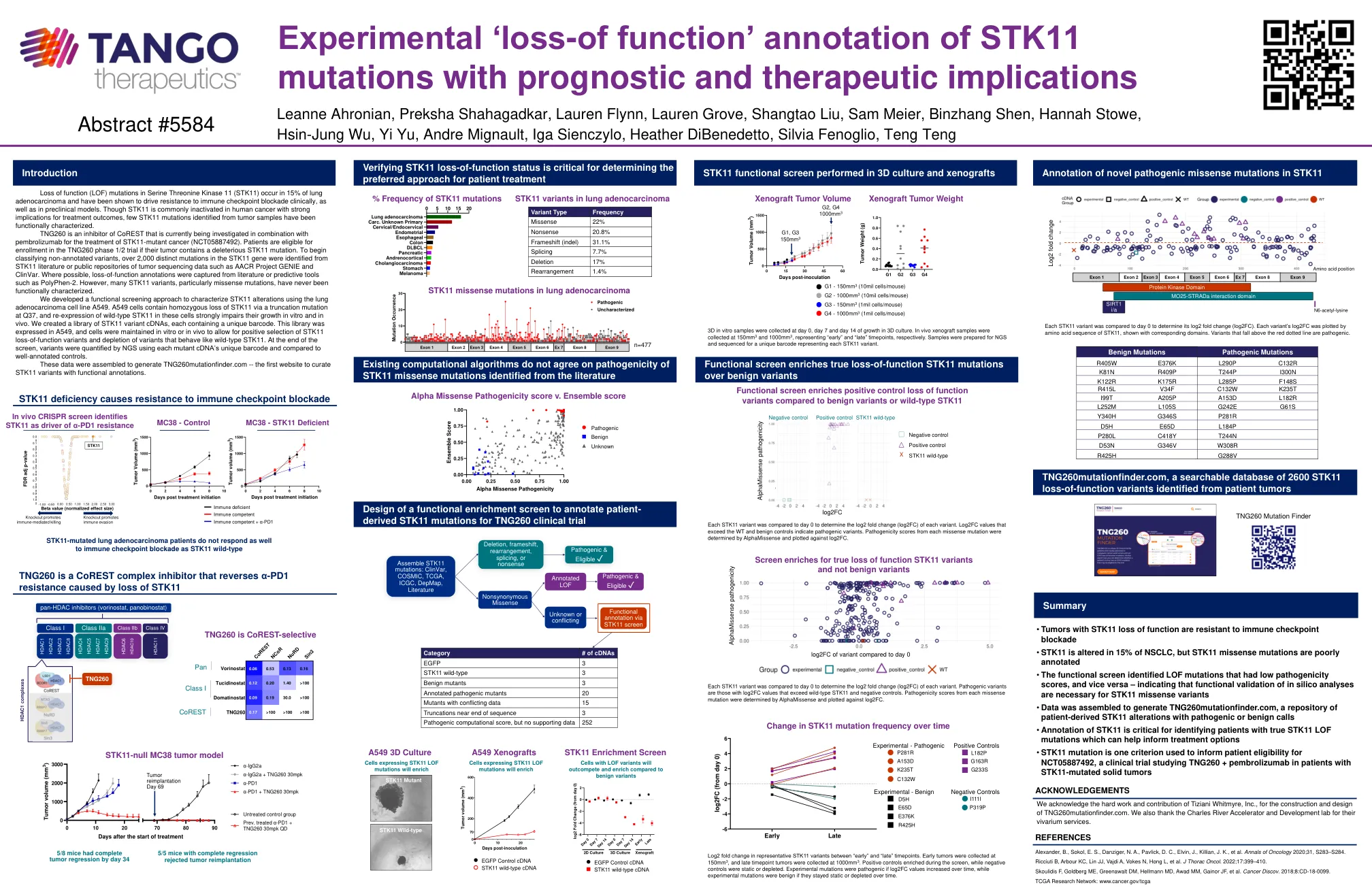
STK11的实验“功能丢失”注释...
丝氨酸苏氨酸激酶11(STK11)中功能(LOF)突变的丧失发生在15%的肺腺癌中,并且已被证明在临床上以及临床前模型中促进了对免疫检查点阻断的抗性。尽管STK11在人类癌症中通常被灭活,对治疗结果的影响很大,但是从功能上表征了从肿瘤样品中鉴定出的STK11突变。TNG260是Corest的一种抑制剂,目前正在研究与Pembrolizumab结合使用STK11-突变癌(NCT05887492)。患者有资格参加TNG260期1/2期试验,如果他们的肿瘤含有有害的STK11突变。为了开始对未经注销的变体进行分类,从STK11文献或肿瘤测序数据的公共存储库中鉴定出超过2,000个不同的突变,例如AACR Project Genie和Clinvar。在可能的情况下,从文献或诸如Polyphen-2之类的预测工具中捕获了功能丧失注释。但是,许多STK11变体,尤其是错义突变,从未在功能上表征。我们开发了一种功能筛选方法,使用肺腺癌细胞系A549表征STK11改变。A549细胞包含通过Q37处的截短突变纯合损失STK11,并且在这些细胞中重新表达了野生型STK11的表达,严重损害了它们在体外和体内的生长。我们创建了一个STK11变体cDNA的库,每个cdnas包含一个唯一的条形码。在屏幕末端,使用每个突变cDNA的独特条形码通过NGS对变体进行了定量,并将其与良好的对照对照进行了比较。该文库在A549中表达,并在体外或体内保持细胞,以允许对STK11功能丧失变体进行积极选择,并且耗尽了像野生型STK11的变体。这些数据被组装成生成TNG260Muntfinder.com-第一个策划具有功能注释的STK11变体的网站。

STK11 是一种潜在的治疗和预后生物标志物,与非小细胞肺癌的免疫浸润相关
目的:本研究旨在阐明丝氨酸苏氨酸激酶 11 (STK11) 在非小细胞肺癌 (NSCLC) 中的作用,特别是其在 KRAS 突变 NSCLC 对抗 PD-1 单克隆抗体治疗的耐药性中的作用。该研究还探讨了 STK11 改变对预后、蛋白质相互作用、免疫细胞参与和药物敏感性的影响。方法:进行全面的生物信息学分析以评估各种 NSCLC 亚型中的 STK11 表达水平和突变谱。该研究将这些发现与临床病理特征相关联,并评估了免疫细胞浸润、免疫微环境和潜在的治疗选择。还进行了分子对接分析以研究与各种抑制剂的相互作用。结果:结果显示整个 NSCLC 中的 STK11 表达升高,突变率为 14%,并且与良好的预后相关。发现 STK11 表达与免疫细胞浸润和以免疫活性较低为特征的冷免疫微环境相关。 Nutlin-3a (-) 被确定为 STK11 突变 NSCLC 病例的潜在治疗选择。分子对接分析提供了与各种抑制剂相互作用的见解,为个性化治疗策略提供了前景。结论:本研究强调 STK11 是 NSCLC 的双重预后和治疗生物标志物。研究结果强调了 STK11 与免疫活动之间的复杂相互作用,为 NSCLC 的个性化治疗方法提供了创新途径。关键词:非小细胞肺癌、STK11、免疫细胞浸润、预后生物标志物、治疗生物标志物、免疫疗法耐药性

依维莫司治疗患有 TSC1、TSC2、NF1、NF2 或 STK11 突变的晚期实体恶性肿瘤患者的 II 期研究
哺乳动物雷帕霉素靶蛋白 (mTOR) 是一种调节细胞生长、增殖和存活的关键蛋白激酶 (1)。mTOR 通路的激活与多种恶性肿瘤的发展有关 (2,3)。这种蛋白激酶主要通过 AKT 和结节性硬化症复合体 (TSC1/TSC2) 的 PI3K 通路激活 (1)。该通路还受肿瘤抑制因子(如 STK11 和 NF1 )的调节,这些因子在不同癌症中经常发生改变 (4,5)。STK11 (也称为 LKB1 )通过激活 AMPK 和磷酸化 TSC2 来激活 mTOR,而 NF1 通过终止 RAS 蛋白的活性状态来阻止下游 mTOR 通路的激活 (4,5)。TSC1、TSC2、STK11 和 NF1 基因突变可导致 mTOR 通路失调并促进肿瘤细胞生长 (6)。因此,抑制 mTOR 可以成为一种治疗携带肿瘤抑制因子(如 STK11、NF1、TSC1 和 TSC2)突变的实体肿瘤的方法。

对患有TSC1,TSC2,NF1,NF2或STK11突变的晚期固体恶性肿瘤患者的Everolimus的II期研究
雷帕霉素(MTOR)哺乳动物靶标是一种关键蛋白激酶,可调节细胞生长,增殖和存活率(1)。 MTOR途径的激活与几种恶性肿瘤的发展有关(2,3)。 该蛋白激酶主要通过AKT和结节性硬化症复合物(TSC1/TSC2)通过PI3K途径激活(1)。 该途径也受肿瘤抑制剂(例如STK11和NF1)的调节,这些途径经常在不同的癌症中改变(4,5)。 stk11,也称为LKB1,通过激活AMPK和TSC2的磷酸化激活MTOR,而NF1通过终止Ras蛋白的活性状态(4,5)来阻止MTOR途径的下游激活。 TSC1,TSC2,STK11和NF1基因中的突变会导致MTOR途径失调并促进肿瘤细胞的生长(6)。 抑制mTOR可以代表一种治疗固体瘤的方法,该方法在STK11,NF1,TSC1和TSC2等肿瘤抑制子中含有突变。哺乳动物靶标是一种关键蛋白激酶,可调节细胞生长,增殖和存活率(1)。MTOR途径的激活与几种恶性肿瘤的发展有关(2,3)。该蛋白激酶主要通过AKT和结节性硬化症复合物(TSC1/TSC2)通过PI3K途径激活(1)。该途径也受肿瘤抑制剂(例如STK11和NF1)的调节,这些途径经常在不同的癌症中改变(4,5)。stk11,也称为LKB1,通过激活AMPK和TSC2的磷酸化激活MTOR,而NF1通过终止Ras蛋白的活性状态(4,5)来阻止MTOR途径的下游激活。TSC1,TSC2,STK11和NF1基因中的突变会导致MTOR途径失调并促进肿瘤细胞的生长(6)。抑制mTOR可以代表一种治疗固体瘤的方法,该方法在STK11,NF1,TSC1和TSC2等肿瘤抑制子中含有突变。
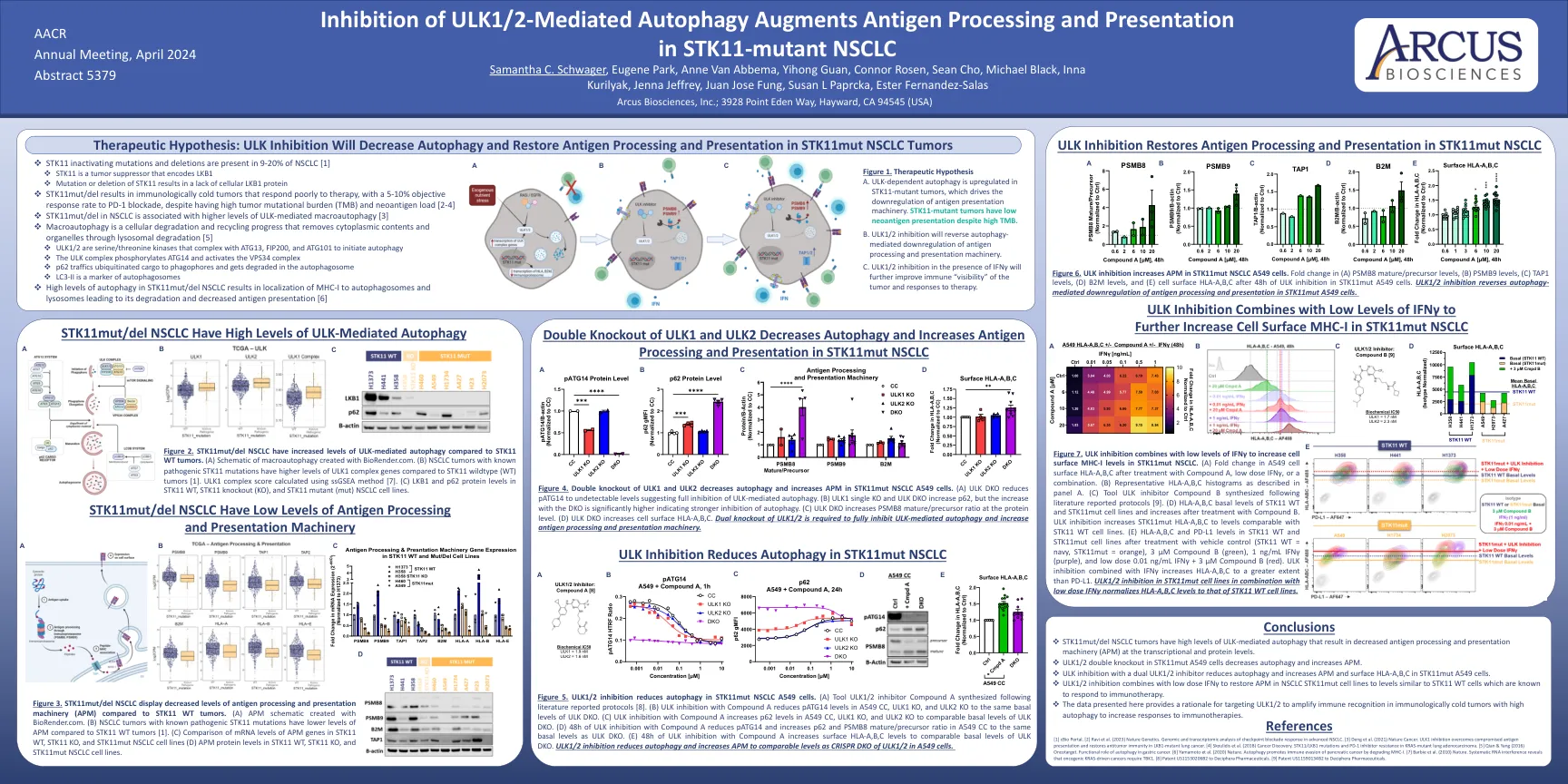
抑制ULK1/2介导的自噬增强抗原加工和表现 ARC-9:一项评估基于伊特鲁马德的治疗组合的随机研究
图2。与STK11 WT肿瘤相比, STK11MUT/DEL NSCLC具有ULK介导的自噬水平升高。 (a)用biorender.com创建的大噬细胞的示意图。 与STK11野生型(WT)肿瘤相比,具有已知致病性STK11突变的NSCLC肿瘤具有更高水平的ULK1复合基因[1]。 ULK1复合分数使用SSGSEA方法计算[7]。 (C)STK11 WT,STK11敲除(KO)和STK11突变体(MUT)NSCLC细胞系中的LKB1和P62蛋白水平。 图4。 ULK1和ULK2的双重敲除可降低自噬并增加STK11MUT NSCLC A549细胞中的APM。 (a)ULK DKO将PATG14降低至无法检测的水平,表明对ULK介导的自噬完全抑制。 (b)ULK1单个KO和ULK DKO增加了p62,但随着DKO的增加,dKO的增加表明自噬抑制更强。 (c)ULK DKO在蛋白质水平上增加了PSMB8成熟/前体比率。 (d)ULK DKO增加了细胞表面HLA-A,b,c。 需要ULK1/2的双重敲除以完全抑制ULK介导的自噬并增加抗原加工和表现机制。STK11MUT/DEL NSCLC具有ULK介导的自噬水平升高。(a)用biorender.com创建的大噬细胞的示意图。与STK11野生型(WT)肿瘤相比,具有已知致病性STK11突变的NSCLC肿瘤具有更高水平的ULK1复合基因[1]。ULK1复合分数使用SSGSEA方法计算[7]。 (C)STK11 WT,STK11敲除(KO)和STK11突变体(MUT)NSCLC细胞系中的LKB1和P62蛋白水平。 图4。 ULK1和ULK2的双重敲除可降低自噬并增加STK11MUT NSCLC A549细胞中的APM。 (a)ULK DKO将PATG14降低至无法检测的水平,表明对ULK介导的自噬完全抑制。 (b)ULK1单个KO和ULK DKO增加了p62,但随着DKO的增加,dKO的增加表明自噬抑制更强。 (c)ULK DKO在蛋白质水平上增加了PSMB8成熟/前体比率。 (d)ULK DKO增加了细胞表面HLA-A,b,c。 需要ULK1/2的双重敲除以完全抑制ULK介导的自噬并增加抗原加工和表现机制。ULK1复合分数使用SSGSEA方法计算[7]。(C)STK11 WT,STK11敲除(KO)和STK11突变体(MUT)NSCLC细胞系中的LKB1和P62蛋白水平。图4。ULK1和ULK2的双重敲除可降低自噬并增加STK11MUT NSCLC A549细胞中的APM。(a)ULK DKO将PATG14降低至无法检测的水平,表明对ULK介导的自噬完全抑制。(b)ULK1单个KO和ULK DKO增加了p62,但随着DKO的增加,dKO的增加表明自噬抑制更强。(c)ULK DKO在蛋白质水平上增加了PSMB8成熟/前体比率。(d)ULK DKO增加了细胞表面HLA-A,b,c。需要ULK1/2的双重敲除以完全抑制ULK介导的自噬并增加抗原加工和表现机制。

CTLA4 阻断可消除 KEAP1/STK11 相关的对 PD-(L)1 抑制剂的耐药性
对于晚期非小细胞肺癌 (NSCLC) 患者,与单独使用 PD-(L)1 抑制剂治疗相比,使用 CTLA4 抑制剂和 PD-1 或 PD-L1 抑制剂(下称 PD-(L)1 抑制剂)的双重免疫检查点阻断 (ICB) 具有更高的抗肿瘤活性和免疫相关毒性。然而,目前还没有经过验证的生物标志物来确定哪些患者会受益于双重 ICB 1,2 。我们在这里表明,在随机 III 期 POSEIDON 试验中,当将 PD-L1 抑制剂 durvalumab 和 CTLA4 抑制剂 tremelimumab 的双重 ICB 添加到化疗中时,患有 STK11 和/或 KEAP1 肿瘤抑制基因突变的 NSCLC 患者可从临床受益,但不能从单独使用 durvalumab 中受益 3 。公正的基因筛选发现,这两种肿瘤抑制基因的缺失是导致 PD-(L)1 抑制耐药的独立驱动因素,并表明 Keap1 的缺失是双重 ICB 疗效的最强基因组预测因子——这一发现在几种 Kras 驱动的 NSCLC 小鼠模型中得到证实。在小鼠模型和患者中,KEAP1 和 STK11 的改变都与不良的肿瘤微环境有关,其特征是抑制性髓系细胞占优势、CD8 + 细胞毒性 T 细胞耗竭,但 CD4 + 效应亚群相对保留。双重 ICB 强效作用于 CD4 + 效应细胞,并将肿瘤髓系细胞区室重新编程为表达诱导型一氧化氮合酶 (iNOS) 的杀瘤表型,其与 CD4 + 和 CD8 + T 细胞一起发挥抗肿瘤功效。这些数据支持使用双重 ICB 化学免疫疗法来减轻患有 STK11 和/或 KEAP1 改变的 NSCLC 患者对 PD-(L)1 抑制的耐药性。

将基因组生物标志物结合起来指导非小细胞肺癌Joris van de haar 1,2,3,Joanne M. Mankor 4,Karlijn Hummelink 5
摘要◥目的:在非小细胞肺癌(NSCLC)中指导免疫检查点阻断(ICB)治疗的STK11,KEAP1和EGFR改变的临床值仍然有争议,因为一些拟议的抗药性生物标志物显示出耐用的ICB反应。这种疾病迫切需要更多特定的组合生物标志物方法。ExperimentalDesign: Todevelopacombinatorialbiomarkerstra- tegy with increased speci fi city for ICB unresponsiveness in NSCLC, we performed a comprehensive analysis of 254 patients with NSCLC treated with ligand programmed death-ligand 1 (PD-L1) blockade monotherapy, including a discovery cohort of 75 patients subjected to whole-genome sequencing (WGS), and独立的验证队列,由169名患者进行肿瘤非正式面板序列。在低(<10 muts / mb)或高(≥10cuts / mb)肿瘤突变负担(TMB)的背景下,评估了STK11 / KEAP1 / EGFR改变的特定级别的ICB反应性。

患者问题:肺癌的创新
ALK (4) NRAS (2) ARID1A (1) NRG1 (2) ATM (2) NTRK (6) ATR (1) P53 (1) BRAF (15) PALB (1) BRCA (2) PDGFR (2) CCND (1) PIK3 (5) CDK4/6 (1) POLD1 (2) CDKN2 (1) POLE (2) ctDNA (3) PRKC (2) CTNNB1 (1) PTCH1 (2) DDR (5) RAF (2) EGFR (10) RAS (2) ERK (2) RET (4) EWSR1 (1) ROS1 (5) EZH2 (1) SMO (1) FBXW7 (1) STK11 (1) FGFR (4) TAF15 (1) GNAQ/11 (2) TMB (4) HRD (7) TP53 (1) IDH (2) TSC (1) KIT (2) UGT1A1 (1) KRAS (24) CCNE1 (1) MDM2 (1) CLDN (3) MEK (3) FET (1) MET (5) FRa (1) MMR (2) FUS (1) MRD (2) HER2 (12) MSI-H (1) MAGE (2) MTAP (4) MUC (1) MYC (1) PD-L1 (12) Nectin-4 (2) PRAME (1) NF1 (6)

基于患者细胞的药物基因组学评估,揭示晚期肺癌的潜在耐药机制和新疗法
实体肿瘤的表达谱。由于 LUAD 在我们的队列中占主导地位(> 80%),我们将 PDC 基因表达谱与 TCGA-LUAD 数据集(n = 230)进行了比较。正如预期的那样,PDC 基因组图谱与肿瘤样本相似,并与正常邻近组织区分开来(图 1B)[36]。PDC 和 TCGA 样本中的组成性体细胞基因突变相似。TP53、RB1 和 BRAF 突变的复发在 PDC 和 TCGA 样本中都得到了高度保留。PDC 中的 EGFR 突变频率较高,而 KRAS、KEAP1 和 STK11 突变的复发低于 TCGA 样本(图 1B)。因此,在 PDC 模型中经常观察到 TP53(49%)、EGFR(31%)和 RB1(8%)的体细胞突变(图 1C)。此外,MET (10%)、CDK4 (6%)、

引用本文:Castillo P、Marginet M、Jares P、García M、Gonzalvo E、Arance A 等。实施 NGS 面板用于临床实践
结果:共对 155 个样本进行了分子分析,但 40 个样本(25.8%)不适合进行测序。在 29 个样本中比较了 BRAF V600 实时聚合酶链反应和靶向 NGS 的临床实用性,观察到非常好的一致性(Kappa = 0.89,95% 置信区间 0.68 ± 1.05)。通过 NGS 在 75 个样本(65%)中发现了致癌突变,其中 53% 是需要个性化治疗的候选人。最常见的突变基因是 BRAF(39%)、TP53(23%)和 NRAS(14%)。其他发生率较低(< 5%)的基因是:PIK3CA、ERBB4、CTNNB1、STK11、FGFR1、SMAD4、KRAS、FGFR3、PTEN 和 AKT。 40% 的样本检测到致癌突变同时发生。在已识别的突变中,TP53 在男性中明显更普遍(男性 31.8% 对比女性 12.2%,P = 0.03),NRAS 在女性中明显更普遍(男性 9.1% 对比女性 24.4%,P = 0.03)。