XiaoMi-AI文件搜索系统
World File Search SystemSox2
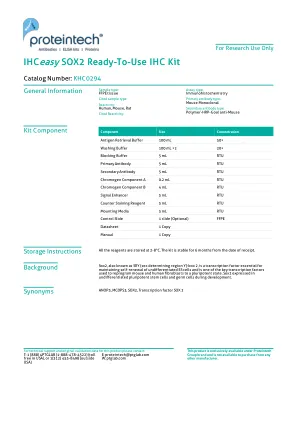
ihceasy sox2 sox2现成的IHC套件
背景Sox2,也称为SRY(性别确定区域Y)-Box 2,是维持未分化的ES细胞自我更新至关重要的转录因子,并且是将小鼠和人成纤维细胞重新编程为多能状态的关键转录因子之一。SOX2在发育过程中在未分化的多能干细胞和生殖细胞中表达。

Oct4、Sox2、Klf4、c-My(OSKM)基因治疗...
雌二醇 [4, 5]。我们小组实施了能够延长雌性大鼠生殖功能的策略。因此,在雌性 MA 大鼠中,我们之前已经证明,从 8 月龄开始对下丘脑内胰岛素样生长因子-I (IGF-I) 进行基因治疗,可将动物的规律性周期延长至 10 个月以上(MA 大鼠停止排卵的年龄),并保持卵巢结构的完整性。在 11 月龄时,接受治疗的大鼠通常表现出保留其规律性的周期性以及正常的卵巢组织学,而对照组在相同年龄时大多无周期性,并且表现出高比例的多囊卵巢和少量黄体 [6]。衰老与表观遗传学动物克隆的发现 [7, 8] 和随后细胞重编程的发展

清理照片的机器学习照片和定量3D神经病理学的表面扫描
图1。SOX2 C-IDR是无序且动态的。a)Sox2的示意图说明了本研究中使用的主要构建体。基于两个不同的预测因子(疾病332(虚线),Alphafold 19归一化PLDDT(实线)),该图显示了障碍预测与残基数的函数。DBD以及广告和富含丝氨酸的区域(有关详细信息,请参见文本)以及带电残基的位置。b)在5 µm浓度下不同SOX2变体的远紫外圆形二分法;全长Sox2(蓝色),C-IDR(灰色),N-DBD(绿色)。光谱是n = 3个独立测量值的平均值。c-d)Sox2荧光标记的单分子转移效率直方图,该荧光标记了DBD的两侧(残基37和120,分子数= 5323)或探测整个C- IDR(残基120-315,分子数量,分子数= 14544)。e)SOX2 C-IDR的荧光寿命分析。2D相关图显示了相对于固有供体荧光(d)的CY3B供体(da)的荧光寿命。动态线基于锯 - 聚合物模型。有关详细信息,请参见文本。f)1 H 15 N-HSQC全长SOX2的频谱。g)全长Sox2(蓝色)的CSCS图。确定DBD(绿色)的 SCSS针对孤立的N-

三聚体BET v 1特异性纳米化引起对IgE结合的强抑制
胚胎干细胞通过形成细菌层具有多能力的潜力和自我恢复能力,从而为胚胎发生提供了主要贡献。这些干细胞多能的保留取决于转录因子的表达/水平,即SOX2,OCT4和NANOG。在器官发生过程中,分子的表达变化也会影响这些干细胞失去多能性并转向谱系选择。随着分化的进展,包括口腔鳞状细胞在内的体细胞的维持也取决于转录因子的差异表达。最近,许多实验性和观察性研究记录了各种人类癌症的致癌作用的重要贡献。在这篇综述中,我们试图总结说明这些主要多能调节剂在口服癌变阶段的推定作用的证据,即口服鳞状细胞癌的起始,进展和预后。

RB613鼠标抗SOX2
BD Horizon Realblue™613(RB613)染料是BD®家族的一部分。这是一个串联荧光色素,在492 nm处具有激发最大(EX MAX),在613 nm处最大发射(EM MAX),使用抗体-DYE结合物测量。由BD®创新驱动,RB613可在光谱和常规的细胞仪上使用,并被蓝色激光器(488-nm)兴奋,并通过561 nm Yellow-Green Laser降低了激发。对于配备蓝色激光器(488-nm)的常规仪器,RB613可用作PE-CF594的替代方案,我们建议使用以610 nm接近的光滤光片(例如,610/20-NM带通滤波器)。用于配备蓝色激光器(488 nm)的光谱仪器,可以与PE-CF594结合使用。RB613平均比蓝色激光器的PE-CF594更明亮。

文章 SOX2 表达和转录活性可识别肉瘤中的癌症干细胞亚群,具有预后意义
摘要:肉瘤的干性由癌症干细胞 (CSC) 中多能性因子(如 SOX2)的表达协调。SOX2 在骨肉瘤中对肿瘤发生和发展的作用已得到很好的研究。然而,SOX2 的促肿瘤发生特性在其他肉瘤亚型中很少得到研究。在这里,我们表明 SOX2 耗竭显著降低了未分化多形性肉瘤 (UPS) 细胞形成肿瘤球和启动肿瘤生长的能力。相反,SOX2 过表达导致体内致瘤性增加。此外,使用允许监测表达 SOX2 和/或 OCT4 的活细胞的报告系统 (SORE6),我们发现 SORE6+ 细胞比 SORE6- 亚群更具致瘤性。与这一发现一致的是,肉瘤患者中的 SOX2 表达与肿瘤等级、分化、侵袭潜力和较低的患者生存率有关。最后,我们研究了一组抗肿瘤药物对 UPS 模型和患者来源的软骨肉瘤系的 SORE6+ 细胞的影响。我们发现,光神霉素类似物 EC-8042 在体外和体内减少 SORE6+ 细胞方面最有效。总体而言,这项研究表明 SOX2 是一种具有肉瘤预后潜力的促肿瘤发生因子。此外,SORE6 转录活性是肉瘤中真正的 CSC 标记,是评估抗肿瘤治疗对 CSC 亚群疗效的极佳生物标记。

NOTCH 和 SOX2 的相互抑制塑造了三阴性乳腺癌中肿瘤细胞的可塑性和治疗逃逸
癌细胞可塑性是三阴性乳腺癌 (TNBC) 化疗和靶向治疗失败的重要原因。治疗诱导的肿瘤细胞可塑性和相关耐药性的分子机制在很大程度上是未知的。使用全基因组 CRISPR-Cas9 筛选,我们研究了用 γ 分泌酶抑制剂 (GSI) 治疗的 NOTCH 驱动的 TNBC 的逃逸机制,并确定 SOX2 是 Notch 抑制耐药性的靶点。我们描述了 Notch 信号和 SOX2 之间的一种新型相互抑制反馈机制。具体而言,Notch 信号通过其 HEY 家族的靶基因抑制 SOX2 表达,而 SOX2 通过与 RBPJ 直接相互作用抑制 Notch 信号。这种机制形成了不同的细胞状态,其中 NOTCH 阳性 TNBC 更像上皮细胞,而 SOX2 表达与上皮-间质转化相关,诱导癌症干细胞特征和 GSI 耐药性。为了抵消单药治疗引起的肿瘤复发,我们分别评估了 GSI-紫杉醇和达沙替尼-紫杉醇联合治疗对 NOTCH 抑制剂敏感和耐药的 TNBC 异种移植的效果。这些独特的预防组合和二线治疗方案依赖于 TNBC 中的 NOTCH1 和 SOX2 表达,能够诱导肿瘤生长控制并减少转移负担。

海马齿状回发育所需的早期SOX2依赖性基因表达程序
海马是认知的大脑区域。人类SOX2转录因子中的突变会导致神经发育缺陷,导致智障和癫痫发作,以及海马发育不良。我们在小鼠中产生了一系列等位基因SOX2条件突变,在不同的发育阶段删除SOX2。SOX2晚期缺失(来自E11.5,通过Nestin-Cre)仅影响产后海马发育;早期的缺失(来自E10.5,EMX1-CRE)显着降低了齿状回(DG),最早的缺失(来自E9.5,FOXG1-CRE)会导致剧烈的异常,几乎完全没有DG。我们识别一组功能相互连接的基因(Gli3,Wnt3a,cxcr4,p73和tbr2),已知在海马胚胎发生中起着重要作用,在SOX2早期突变体中被下调,以及(Gli3和cxcr4)直接通过SOX2键入SOX2;它们的下调提供了导致缺陷的合理分子机制。对EMX1-CRE小鼠模型的电生理研究显示CA1和CA3区域的兴奋性传播改变了。对EMX1-CRE小鼠模型的电生理研究显示CA1和CA3区域的兴奋性传播改变了。

AKT1 作为铂类耐药 SOX2 阳性卵巢癌细胞的治疗靶点
卵巢癌仍然是最致命的妇科恶性肿瘤,主要是因为其化疗耐药性和高复发率。越来越多的证据表明,SOX2 异常表达与卵巢癌患者的化疗耐药性和不良预后有关。在本研究中,我们旨在阐明卵巢癌细胞中 SOX2 异常表达的机制。通过检查多种卵巢癌细胞系和一组临床肿瘤样本,我们观察到卵巢癌细胞系和肿瘤中 SOX2 广泛过表达。为了确定导致卵巢癌细胞中 SOX2 过表达的信号通路,我们筛选了一组针对 30 种主要细胞激酶的小分子激酶抑制剂。在确定的热门药物中,AKT 抑制剂是其中之一。我们证明,抑制或敲低 AKT1 可大幅下调 SOX2 蛋白水平,损害 SOX2 阳性卵巢癌细胞的生长和干细胞特性,并显著提高 SOX2 阳性卵巢癌细胞对铂类药物的敏感性。从机制上讲,我们发现 AKT1 主要通过增强其蛋白质稳定性来驱动 SOX2 过表达,并通过在 T116 位点磷酸化 SOX2 来实现这一点。总之,我们的研究揭示了卵巢癌中驱动 SOX2 过表达的潜在机制,并强调了药理学抑制 AKT1 是一种潜在的治疗策略,可提高 SOX2 阳性卵巢癌对铂类药物的敏感性。

SOX2,OCT4和NANOG:口服致癌作用中的核心胚胎干细胞多能调节剂
摘要:在这项工作中,使用简单的溶剂热技术制备了UIO-66-NH 2 /GO纳米复合材料,并使用现场发射扫描电子显微镜(FE-SEM),能量分散性的X射线光谱镜(EDS)和X射线散布(X-Ray衍射(XRD)对其结构和形态进行了表征。提出了一种用于检测表蛋白(EP)的增强的电化学传感器,该传感器利用UIO-66-NH 2 /GO纳米复合材料修饰的筛网印刷石墨电极(UIO-66- NH 2 /GO /SPGE)。制备的UIO-66-NH 2 /GO纳米复合材料改善了SPGE对EP的氧化还原反应的电化学性能。在优化的实验条件下,该传感器显示出明显的检测限制(LOD)为0.003 µm,线性动态范围为0.008至200.0 µm,提供了一个高功能的传感EP平台。此外,利用差分脉冲伏安法(DPV)研究了在UIO-66-NH 2 /GO /SPGE表面上研究EP和拓扑替康(TP)(TP)的同时进行电催化的氧化。DPV测量结果表明存在EP和TP的两个明显的氧化峰,峰电势分离为200 mV。最后,在药物注射中,成功使用了UIO-66-NH 2 /GO /SPGE传感器来对EP和TP进行定量分析,从而产生了高度令人满意的结果。