XiaoMi-AI文件搜索系统
World File Search SystemSox2

基于患者来源的人类成纤维细胞的戈谢病体外药物筛选模型的表征
图 1. 人类 iPSC 发育成脑类器官的代表性图像。(A) 健康个体的人类诱导多能干细胞集落。(B) 接种在圆底板中的 DIV2 胚状体。(C) DIV7 胚状体,神经外胚层的形成表现为胚状体周围的光晕。(D) DIV10 胚状体嵌入 Matrigel 中,支持神经上皮的形成。(E) DIV13 脑类器官,神经花结的存在代表干细胞分化为神经祖细胞。(F) DIV34 脑类器官,其中类器官的直径 >1,000 µm。(G) DIV35 类器官中神经元前体 SOX2(红色)和成熟神经元 NeuN(白色)的免疫组织化学染色。
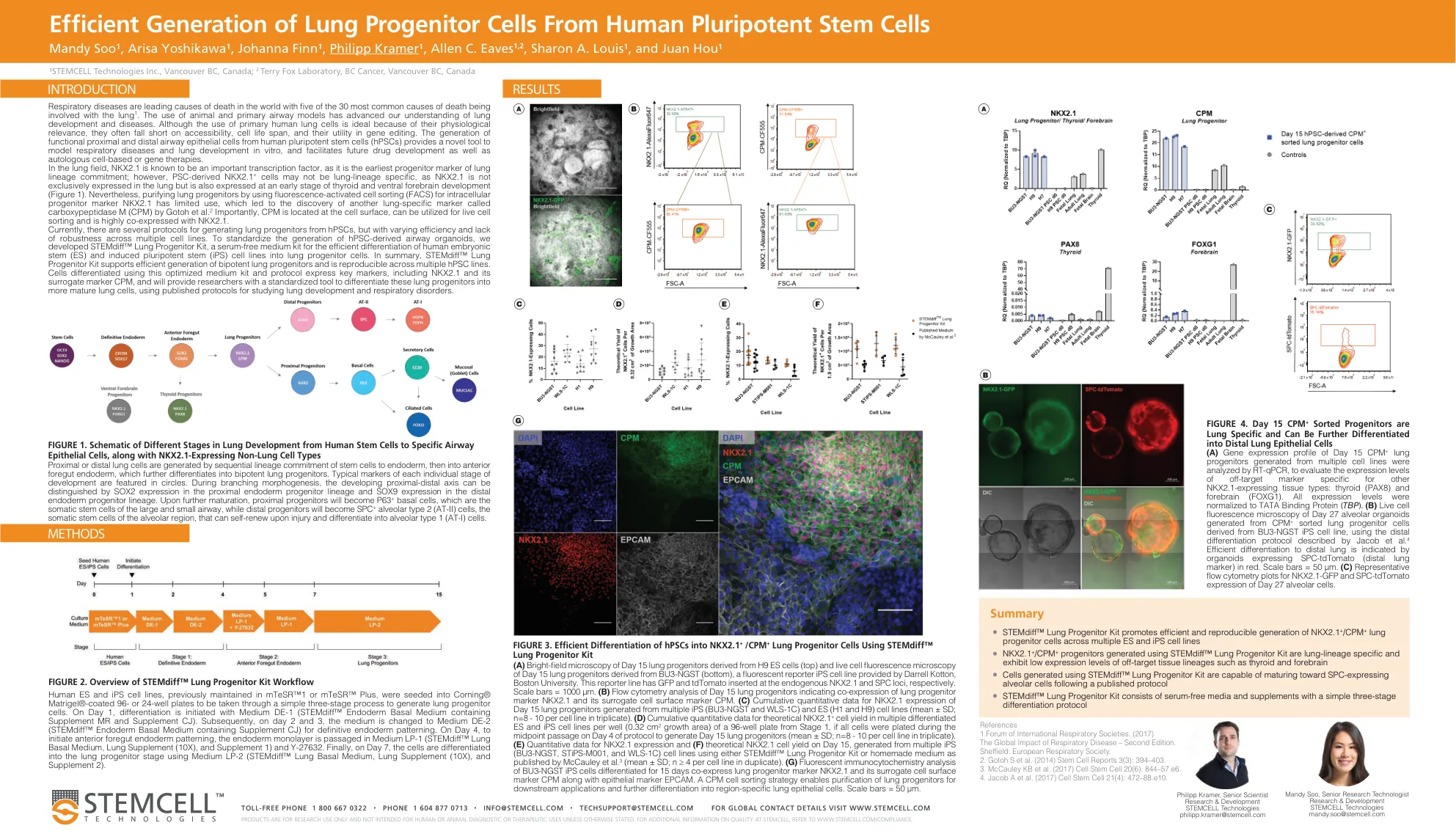
SP00248-从人类多能干细胞高效生成肺祖细胞
近端或远端肺细胞是由干细胞按顺序谱系分化到内胚层,然后进入前肠内胚层,进一步分化为双能肺祖细胞而产生的。每个发育阶段的典型标记以圆圈表示。在分支形态形成过程中,可以通过近端内胚层祖细胞谱系中的 SOX2 表达和远端内胚层祖细胞谱系中的 SOX9 表达来区分发育中的近端-远端轴。进一步成熟后,近端祖细胞将变成 P63 + 基底细胞,即大气道和小气道的体细胞干细胞,而远端祖细胞将变成 SPC + 肺泡 2 型 (AT-II) 细胞,即肺泡区域的体细胞干细胞,它们在受伤后可以自我更新并分化为肺泡 1 型 (AT-I) 细胞。

crispr.pdf的生成
使用表达人OCT4,KLF4的综合质粒对健康的24岁女供体的人类皮肤成纤维细胞进行了重编程; L-MYC(OSKM),SOX2和LIN28(Okita等,2011),并进行了多个段落。然后使用基于蛋白质的CRISPR/CAS9基因组编辑方法来生成纯合的KIF1C基因敲除线(资源表)生成纯合的KIF1C敲除线(资源表)。首先,IPSC-CO用KIF1C基因的两个核糖核蛋白(RNP)复合物核成核成核。荧光标记的tracrrna(atto550)可用于选择通过荧光激活的细胞分选(FACS)成功地培养RNP复合物的细胞。然后,在单细胞播种后,手动采摘菌落,通过PCR筛选并扩展了几段段落。。

MACC1调节LGR5以促进结直肠癌的癌症干细胞特性
从HIV-1 + 2,乙型肝炎和丙型肝炎中分离出外周血单核细胞(PBMC)。pBMC,包括人类基因Oct3/4,Sox2,c-Myc和klf4的矢量。HIPSC线BIHI292-A源自单个菌落,并在E8培养基中保持未分化的HIPSC的典型形态(图1 a)。通过PCR确认缺乏仙台病毒载体(Suppl。图1 a)。BIHI292-A HIPSCS ECT3/4,SSEA-4,NANOG和TRA-1 - 60作为未分化HIPSC状态的典型标记,如使用免疫细胞化学所示(图1 b)。进一步的流式细胞仪证实了OCT3/4,SSEA-4,NANOG和TRA-1-60表达在超过96%的BIHI292-A HIPSC中的SSEA-4,Nanog和Tra-1-60表达中的干性标记表达(图。1 c)。g带核分型在GTG上进行(使用GIEMSA的胰蛋白酶G带)进行染色的中期染色体,并揭示了正常的雌性Karyo 46型,XX(图1 D)。 单核苷酸多态性分析表明,与患者的PBMC相比,BIHI292-A HIPSC线没有任何拷贝数变量> 2 Mb> 2 MB> 5 MB(表1)。 短串联重复(STR)分析的结果表明,BIHI292-A细胞系和患者的PBMC的遗传认同是相同的(表1)。 Sanger测序证实了两个Exon 2中的两个Trem2杂合突变C.313del(P.Ala105fs)和C.199del(p。is67fs)(图) 1 e)。 图1 D)。单核苷酸多态性分析表明,与患者的PBMC相比,BIHI292-A HIPSC线没有任何拷贝数变量> 2 Mb> 2 MB> 5 MB(表1)。短串联重复(STR)分析的结果表明,BIHI292-A细胞系和患者的PBMC的遗传认同是相同的(表1)。Sanger测序证实了两个Exon 2中的两个Trem2杂合突变C.313del(P.Ala105fs)和C.199del(p。is67fs)(图1 e)。图为了确认患者突变的存在(Buthut等,2023),在TREM2基因的外显子2中为复合杂合突变进行了BIHI292-A HIPSC的测序。通过将分化为三个细菌层的细胞进行分化,测试了多能分化势。分化测试证实,BIHI292-A HIPSC具有分化为内胚层(CD184 +,SOX17 +),Meso Dermal(CD140B +,CD144 +)和外胚层(PAX-6 +,SOX2 +)细胞的潜力(1 f)。BIHI292-A HIPSC对支原体进行了阴性测试(Suppl。1 b)。
8C 样细胞在体外捕获人类合子基因组激活程序
胚胎基因组的激活标志着发育生物体中第一次主要的转录浪潮。小鼠 2 细胞胚胎和人类 8 细胞胚胎中的合子基因组激活 (ZGA) 对发育至关重要。本文,我们报告在幼稚胚胎干细胞中发现了人类 8 细胞样细胞 (8CLC),其转录类似于人类 8 细胞胚胎。它们表达 ZGA 标记,包括 ZSCAN4 和 LEUTX ,以及转座因子,例如 HERVL 和 MLT2A1 。8CLC 显示降低的 SOX2 水平,可在体外使用 TPRX1 和 H3.Y 标记蛋白进行识别。转录因子 DUX4 的过表达和剪接体抑制会增加人类 ZGA 样转录。令人兴奋的是,8CLC 标记物 TPRX1 和 H3.Y 也在 ZGA 阶段的 8 细胞人类胚胎中表达,因此可能与体内相关。 8CLCs 为表征人类 ZGA 样转录提供了独特的机会,并可能为人类胚胎发生早期事件提供重要见解。

使用靶向和全基因组测序对 SCLC 患者的循环游离 DNA 进行分析
SCLC 肿瘤的基因组分析已发现大量拷贝数变异 (CNA) 和高突变率。 4、5 肿瘤抑制基因肿瘤蛋白 p53 基因( TP53 )和视网膜母细胞瘤 1 基因( RB1 )几乎普遍失活和普遍丢失,转录调节因子(SRY-box 转录因子 2 基因 [ SOX2 ]、核因子 IB 基因 [ NFIB ]、v-myc 禽类粒细胞瘤病毒致癌基因肺癌衍生同源基因 [ MYCL1 ]、v-myc 禽类粒细胞瘤病毒致癌基因神经母细胞瘤衍生同源基因 [ MYCN ] 和 v-myc 禽类粒细胞瘤病毒致癌基因同源基因 [ MYC ])频繁扩增,Notch 和组蛋白乙酰转移酶基因(CREB 结合蛋白基因 [ CREBBP ] 和 E1A 结合蛋白 p300 基因 [ EP300 ])反复突变。 4-6 肿瘤活检,尤其是连续活检,对于 SCLC 来说是一项重大挑战,7,8

用壳测量
图4。刺激记录和使用壳测量。(a)带有封装器官的3D壳MEA的图像。在孵化器内部保留的同时,刺激和记录了类器官。(b)3D-Shell MEA的示意图,并标有北,东和西的三个传单。(c)图显示了通过所有三个电极将20 µA的刺激电流发送到类器官时,显示了记录的电压。(d)所有三个电极的记录电压轮廓图显示从类器官收集的信号。与(c)中所示的刺激相对应的峰将从此轮廓中删除。(e)八周龄的类器官的代表性最大强度Z练习图显示了核(I)和绿色,紫色,紫色和黄色(ii)中所示的核(Hoechst),神经元干细胞(SOX2)和轴突(NF-H)的存在。染色说明了器官内的细胞同质性。在20倍拍摄图像。比例尺为100 µm。

干细胞研究-DEF -LAB
独特的干细胞线标识符BCHI007-A(HNDS0005-01 #B)BCHI007-A-1(HNDS0005-01#B2 + / +)BCHI009-A(HNDS0002-01 #D) BCHi011-A-1 (HNDS0003-01 #F CC39 + / + ) Alternative name(s) of stem cell lines HNDS0005-01 #B HNDS0005-01 #B2 + / + HNDS0002-01 #D HNDS0002-01 #D CC26 + / + HNDS0003-01 #F HNDS0003-01 #F CC39 + / + Institution波士顿儿童医院的联系信息wardiya.afsharsaber@childrens.harvard.edu; Mustafa.sahin@childrens.harvard.edu类型的细胞系IPSCS来源的人类ESC或IPSC BCHI007-A(HNDS0005-01 #B)所需的其他起源信息,年龄:21,性别:性别:f,f,f,种族:白人:白色; BCHI009-A(HNDS0002-01 #D),年龄:20,性别:M,种族:白色; BCHI011-A(HNDS0003-01 #F),年龄:4,性别:M,种族:白色。细胞源成纤维细胞克隆性克隆细胞方法的重编程仙台病毒,非整合(OCT4,SOX2,KLF4和HC-MYC)遗传修饰是

NeuroDisk:一种自动化神经成像遗传学中连续询问驱动的发现的AI方法
图4。刺激记录和使用壳测量。(a)带有封装器官的3D壳MEA的图像。在孵化器内部保留的同时,刺激和记录了类器官。(b)3D-Shell MEA的示意图,并标有北,东和西的三个传单。(c)图显示了通过所有三个电极将20 µA的刺激电流发送到类器官时,显示了记录的电压。(d)所有三个电极的记录电压轮廓图显示从类器官收集的信号。与(c)中所示的刺激相对应的峰将从此轮廓中删除。(e)八周龄的类器官的代表性最大强度Z练习图显示了核(I)和绿色,紫色,紫色和黄色(ii)中所示的核(Hoechst),神经元干细胞(SOX2)和轴突(NF-H)的存在。染色说明了器官内的细胞同质性。在20倍拍摄图像。比例尺为100 µm。

合成和基因组调控元件揭示小鼠胚胎干细胞中顺式调控语法的一些方面
摘要 在胚胎干细胞 (ESC) 中,核心转录因子 (TF) 网络建立了多能性所必需的基因表达程序。为了解决四种关键 TF 之间的相互作用如何促进小鼠 ESC 中的顺式调控,我们分析了两个由 SOX2、POU5F1 (OCT4)、KLF4 和 ESRRB 的结合位点组成的大规模并行报告分析 (MPRA) 文库。合成的顺式调控元件与具有可比结合位点配置的基因组序列之间的比较揭示了调控语法的某些方面。合成元件的表达受结合位点的数量和排列的影响。这种语法对基因组序列的作用很小,因为基因组序列的相对活性最好通过预测的结合位点占用率来解释,而与结合位点身份和定位无关。我们的结果表明,转录因子结合位点 (TFBS) 的影响受位点顺序和方向的影响,但在基因组中,TF 的整体占用率是活性的主要决定因素。