XiaoMi-AI文件搜索系统
World File Search SystemTDTOMATO

新型转基因斑马鱼系用于研究 CHRNA3‐...
摘要 乙酰胆碱 (ACh) 是周围神经系统 (PNS) 和中枢神经系统 (CNS) 的重要神经递质,它通过烟碱型乙酰胆碱受体 (nAChR) 和毒蕈碱型乙酰胆碱受体 (mAChR) 发出信号。在这里,我们探讨了三个 nAChR 亚基 chrna3 、 chrnb4 和 chrna5 的表达模式,它们位于进化保守的簇中。在多种脊椎动物中,这种紧密的基因组定位可能表明共同功能和/或共同表达。通过新型转基因斑马鱼系,我们观察到 PNS 和 CNS 内广泛表达。在 PNS 中,我们观察到 chrna3 tdTomato 、chrnb4 eGFP 和 chrna5 tdTomato 在肠道神经系统中的表达; chrna5 tdTomato 和 chrnb4 eGFP 位于侧线的感觉神经节中;而 chrnb4 eGFP 位于耳朵中。在中枢神经系统中,chrnb4 eGFP 和 chrna5 tdTomato 的表达出现在视网膜中,这三种基因均在大脑的不同区域表达,其中一部分 chrna3 tdTomato 和 chrnb4 eGFP 细胞被发现是投射到侧线的抑制性传出神经元。在脊髓内,我们在运动网络内识别出表达 chrna3 tdTomato、chrnb4 eGFP 和 chrna5 tdTomato 的不同神经元群,包括表达 dmrt3a 的中间神经元和表达 mnx1 的运动神经元。值得注意的是,每个半节段的三到四个初级运动神经元均被 chrna3 tdTomato 和 chrnb4 eGFP 标记。有趣的是,我们在每个半节段中发现了一个 sl 型次级运动神经元,该神经元强烈表达 chrna5 tdTomato 并同时表达 chrnb4 eGFP。这些转基因系为 nAChRs 在运动网络中的潜在作用提供了见解,并为探索它们在整个神经系统一系列组织中尼古丁暴露和成瘾的作用开辟了途径。

沉默基因可以降低胆固醇
在 Hepa 1-6 Pcsk9 tdTomato 细胞中进行的体外筛选表明,基于 ZFP 的 ETR 是 Pcsk9 表观沉默的最有效平台。a,左上角,用于比较 Hepa 1-6 Pcsk9 tdTomato 细胞系中不同 ETR 平台效率的实验程序图。右上角,Hepa 1-6 Pcsk9 tdTomato 细胞系的示意图,其中 TAV2A- tdTomato 盒式磁带以框架内的方式靶向 Pcsk9 的最后一个外显子。右下角,不同 ETR 平台的示意图,显示它们与 Pcsk9 启动子区域的 CGI 的相对结合。图片来源:Nature (2024)。DOI: 10.1038/s41586-024-07087-8

13159黄金
抽象的骨髓基质/干细胞代表了一个静止的细胞群,该细胞种群随着年龄的增长和响应损伤,维持骨骼质量和修复而补充成骨细胞骨形成细胞库。在体内体外和骨形成的基质/干细胞分化的有效介质是物理负荷,但仍不清楚负载诱导的骨形成是否需要对这些常驻基质/干细胞的成骨分化。因此,在这项研究中,我们利用瘦素受体(LEPR)来识别和追踪骨髓基质细胞对体内骨骼的机械加热的贡献。十二周龄的LEPR-CRE; TDTOMATO小鼠接受以11 n峰值负载的压缩胫骨负载,用于40个循环,每隔一天,每天持续2周。组织学分析表明,LEPR-CRE; TDTOMATO +细胞在血管周围围绕围绕骨骼出现,并将骨表面填充为衬里细胞或成骨细胞,然后再经历骨细胞生成。lepr-cre; tdtomato +基质细胞在骨髓中随着年龄的增长而增加,但不遵循胫骨压缩负荷的应用。机械载荷会引起骨骼质量和骨骼锻炼参数的增加,但不会引起LEPR-CRE的增加; TDTOMATO +成骨细胞或成骨细胞。为了研究LEPR细胞中的腺苷酸环化酶6(AC6)是否有助于这种机械适应性反应,LEPR-CRE; TDTOMATO小鼠被进一步交叉
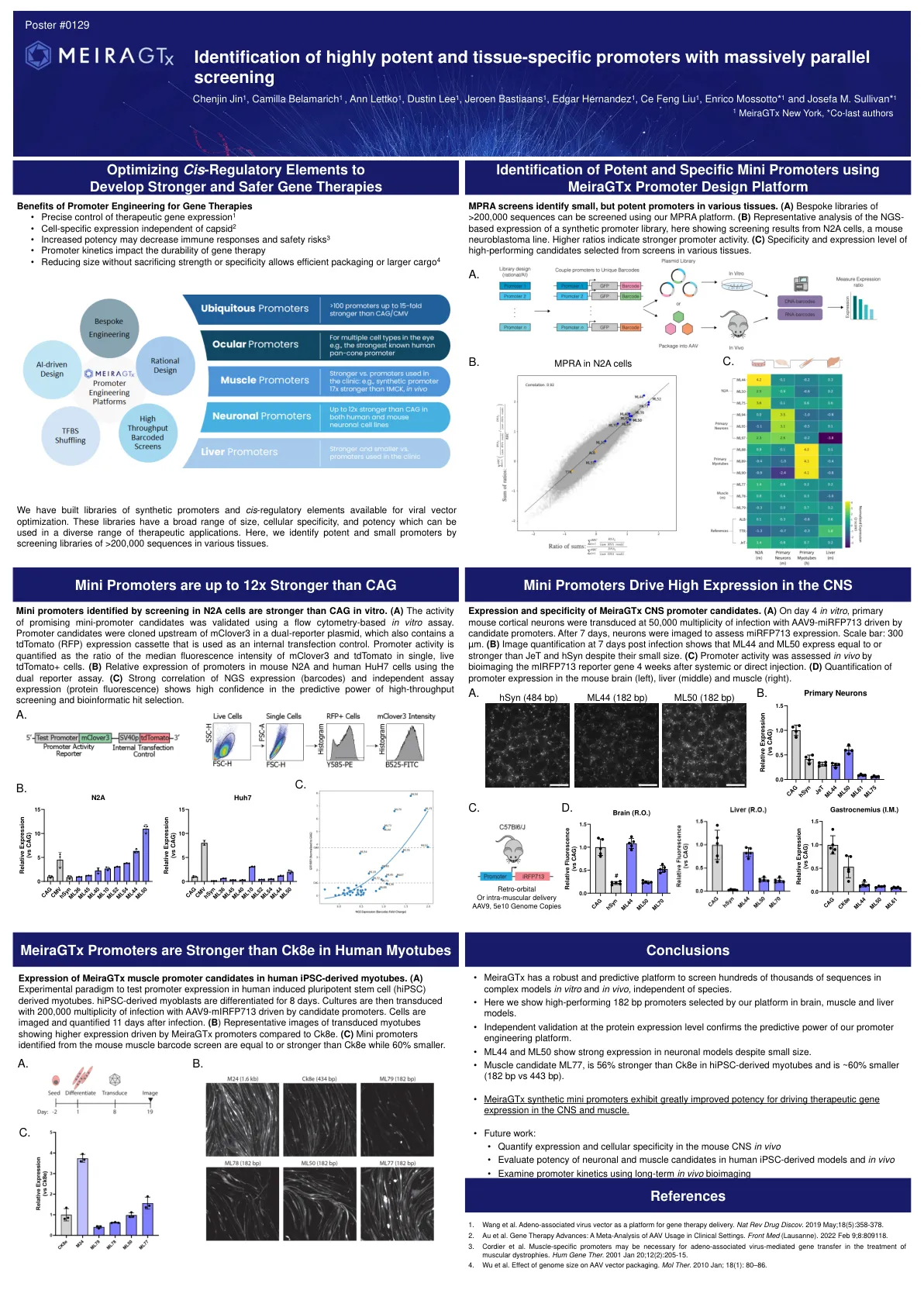
鉴定具有大量平行筛选的高效和组织特异性启动子
迷你启动子在体外比CAG强。(a)使用基于流式细胞术的体外测定法对有希望的迷你启动候选者的活性进行了验证。启动子候选物被克隆在双重孢子质粒中的McLover3上游,该质粒还包含TDTOMATO(RFP)表达盒,该盒被用作内部转染对照。启动子活性被量化为单个活的TDTOMATO+细胞中McLover3和TDTomato的中位荧光强度的比率。(b)使用双报告基因测定法分析,启动子在小鼠N2a和人HuH7细胞中的相对表达。(c)NGS表达(条形码)和独立测定表达(蛋白质荧光)的强相关性表现出对高通量筛选和生物信息学命中选择的预测能力的高信心。

对小鼠海马进行单核 RNA 测序,以评估体内基因编辑
图 3. (A) 小鼠 1 海马的细胞类型识别和 tdTomato 报告基因转录水平 (A) 小鼠 1 的带注释的综合 UMAP 投影显示实验和对照条件下存在 12 种不同的细胞类型。簇 0 和簇 12 被识别为阳性对照成纤维细胞刺突。

小鼠体内编辑肺干细胞以实现持久的基因校正
体内基因组校正有望产生持久的疾病治疗方法;然而,有效的干细胞编辑仍然具有挑战性。在这项研究中,我们证明优化的肺靶向脂质纳米颗粒 (LNP) 能够在干细胞中进行高水平的基因组编辑,从而产生持久的反应。在可激活的 tdTomato 小鼠中静脉注射基因编辑 LNP 可实现 >70% 的肺干细胞编辑,并在 >80% 的肺上皮细胞中维持 tdTomato 表达 660 天。解决囊性纤维化 (CF),NG-ABE8e 信使 RNA (mRNA) – sgR553X LNPs 介导 >95% 的囊性纤维化跨膜传导调节器 (CFTR) DNA 校正,恢复原发性患者支气管上皮细胞中的 CFTR 功能,相当于 Trikafta 治疗 F508del,校正肠道类器官并校正 CF 小鼠 50% 肺干细胞中的 R553X 无义突变。这些发现引入了 LNP 支持的组织干细胞编辑,用于疾病修饰基因组校正。G

使用笼子乙基雌二醇和诱导的Cre/Lox系统迈向眼睛的光介导的基因疗法
摘要:越来越多地,正在用病毒介导的基因疗法治疗视网膜病理。能够以光特定针对视网膜病理区域的病毒转基因表达,我们确定了视网膜组织的体内光活化基因表达para-digm。基于诱导型Cre/Lox系统,我们发现乙基雌二醇是他莫昔芬的合适替代品,因为乙基雌二醇更适合通过光敏保护化合物(即“笼子”)修饰。在计算机结合研究中支持了突变的人雌二醇受体的乙基甲基二醇作为配体的鉴定,该研究表明笼中乙基雌二醇的结合降低。用依赖性的TDTOMATO报告基因转基因注入双转基因GFAP-CREERT2小鼠的眼睛中,然后用450 nm的光照射。光活化显着增加了视网膜TDTOMATO表达。因此,我们展示了为眼睛开发靶向的,光介导的基因治疗的第一步。

nocceptin配体的发育和成人纹状体模式标记具有多巴胺投影的脑膜体种群
纹状体多巴胺信号传导。使用前摄取蛋白 - 碳报告小鼠系列,我们表征了小鼠背纹状体中PNOC mRNA表达的高度选择性的脑膜图模式,反映了PNOC的早期发育表达。在腹侧纹状体中,将PNOC表达聚集在伏隔核和内侧壳中,包括成年纹状体。我们发现PNOC TDTOMATO报告基因细胞在很大程度上包括多巴胺受体D1(DRD1)表达培养基的棘突投射神经元,位于背纹状体中,已知在
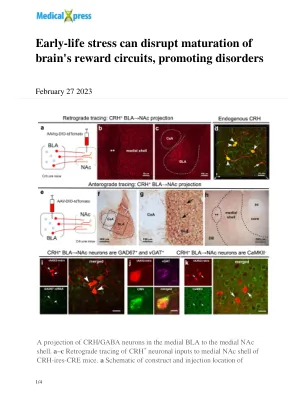
早期的压力会破坏大脑奖励电路的成熟,促进疾病
AAV2 -RETRO -CAG -FLEX -TDOMATO -WPRE病毒,可允许逆行进入投影神经元,从而为NAC提供传入的输入。b示例局部感染的CRH +轴突末端的共聚焦显微照片在内侧NAC壳中。c逆行跟踪将内侧BLA识别为CRH + NAC输入的强大来源。d 3D图像(z-stack;0.5μm步长)确认在AAV-RETRO感染细胞(红色)的BLA中定位,共表达内源性CRH(绿色);双重标记的神经元=黄色。e - 从BLA到内侧NAC壳的CRH +轴突投影的g顺行追踪。e,AAV1-DIO-TDTOMATO构建体和病毒遗传实验设计。f病毒注射仅限于中央杏仁核(CEA)的BLA,G,通过在BLA CRH +神经元中的TDTomato选择性表达显示。h bla-origin CRH +轴突和内侧NAC壳中的端子。i - K病毒注射到BLA中的内侧NAC壳逆转录感染的SOMATA。i将荧光原位杂交(FISH)与BLA中CRH +细胞中的GAD67 mRNA进行免疫染色。箭头指向共定位的GAD67 mRNA和病毒 - 重复蛋白标记。j a bla→NAC细胞(红色)共表达内源性CRH(绿色)和VGAT(Magenta),但K不会共表达谷氨酸能标记Camkii。** = Calleja的主要岛,AC前委员会,DB对角线带。i和k中的比例尺= 10 µm。在至少两个独立的垃圾中评估了在小鼠中评估发现发现,病毒注射,投射评估和免疫组织化学的。信用:自然通讯(2023)。doi:10.1038/s41467-023-36780-x
通过原位转化的脑室区域神经干细胞的原位转化
This protocol describes the surgical procedure for co-electroporation of two plasmids targeting neu- ral stem cells (NSCs) in the lateral ventricle of mouse postnatal day 2 (P2) pups: a nonintegrating plasmid encoding for the piggyBase transposase and Cas9 and an integrating piggyBac vector car- rying the oncogenes, CRISPR guide RNAs and a TDTOMATO荧光报告蛋白通过倒末端重复序列(ITRS)倾斜(图1)。在电穿孔后,瞬时CAS9表达会导致肿瘤抑制基因失活,而PiggyBase介导的PIG-GYBAC货物的整合确保了靶向NSC及其后代中的癌基因和流动性记者的稳定表达。的整合是由PiggyBase转疗的酶促活性介导的,该转移的酶活性通过切割和粘贴机制在受体细胞基因组中的TTAA位点识别并将其与它们的内容一起插入。NSC的靶向是通过最小的人GFAP(HGFAPMIN)启动子序列1-3驾驶PiggyBase/cas9的驱动表达来实现的。