XiaoMi-AI文件搜索系统
World File Search SystemTH17

基于肠道环境的前所未有的肥胖糖尿病治疗
它不能控制它。一些特定的细菌控制肠道中免疫细胞的质量和数量。肠道最初是一种维持抗炎的器官,并且控制着特别居住的肠道免疫的已知肠道细菌之一是SFB(分段的丝状细菌)。 SFB是一种非常独特的细菌,可在肠上皮细胞中定殖,并使用一种称为伴侣(微生物粘附触发的内吞作用)的方法将抗原传递到肠粘膜中的T细胞中,并诱导具有抗原特异性抗激发性抗炎特性的TH17细胞,以替代小肠。众所周知,Th17细胞的性质不同,取决于它们诱导的细菌和诱导的位置,SFB诱导的Th17细胞具有抗炎并增强肠壁。尽管SFB-TH17细胞在全身糖和能量代谢中的作用尚不清楚,但我们发现SFB-TH17细胞具有抗肥胖和糖尿病的作用,并报道高糖/高蔗糖破坏其维持机制2)。有趣的是,发现在SFB单殖民化小鼠中不会发生高糖引起的SFB减少,该小鼠仅在无菌环境中建立了SFB,并且是依赖于SFB以外其他肠道细菌的机制。在高蔗糖和高蔗糖水负荷的情况下,我们集中在一种称为FROD的物种(粪便脂质啮齿动物)上,这是由于高蔗糖而导致的最大变化,并进行了一个SFB和FROD,在无菌小鼠中,SFB和FROD在较高的小鼠中得到了群体的群体。小肠Th17细胞被打破。据说这种机制会导致高蔗糖分解肠道细菌和肠道免疫的稳态维持机制。 最好的糖尿病治疗尚未确定。稳态Th17细胞还保持其功能,而不会损害其稳态至一定的蔗糖浓度,这表明最佳蔗糖浓度有阈值。将来,希望设定可以帮助人们保持健康,维持肠道细菌和肠道免疫的适当摄入量,并检查可以在这种环境下维持肠道免疫稳态的益生菌将成为克服肥胖和糖尿病的治疗策略。

乌司他丁 - Turkderm-Turk Arch Dermatol Venereol 2022
自然杀伤 (NK) 和 CD4 + T 细胞分化为 CD4 + Th1 表型。IL-23 是一种含有 p40 亚基的异二聚体蛋白质,p40 亚基通过二硫键与 p19 亚基相连。IL-23 通过激活 IL-6 和 TGF-b 来诱导 Th17 细胞与 TNF- α 的产生,从而发挥作用。IL-12 和 IL-23 的 p40 亚基均与 IL-12 受体- β 1 (IL-12R β 1) 结合。IL-12p35 和 IL-23p19 亚基分别与 IL-12R β 2 和 IL-23R 结合。这样,IL-12 和 IL-23 尽管在结构上有一些相似之处,但它们控制着不同的免疫途径。通过刺激 CD4 + T 细胞分化为 CD4 + Th1 表型,IL-12 促进 IFN- γ 、TNF- α 和 IL-2 的分泌。相反,IL-23 与 IL-21 和 TGF- β 结合,导致 CD4 + Th17 细胞分化,从而导致其他细胞因子的分泌,例如 L-17、IL-22、TNF- α 和 IL-1 β 。乌司奴单抗是一种抑制 p40 蛋白的人重组 G1 (lgG1) 单克隆抗体。它通过这种 p40 抑制来发挥作用,从而阻断 IL-12 和 IL-23 3,4 。

IL-17 and-il-22-as-as-as-stable-...
IL-17和IL-22之间的相互作用对于维持组织稳态至关重要[9]。在发现Th17细胞作为T-Helper细胞的独立线[10,11]后,它们在炎症中的作用变得更加明显,它分泌IL-17和IL-22细胞因子。仍然需要澄清其分泌的刺激,但是在人类中,与小鼠模型不同,它们并不总是协同分泌的[12,13]。通常,IL-17主要是一种炎性细胞因子,通过增强抗菌肽,粒细胞和中性粒细胞在周围组织中积累的上皮上皮释放来预防感染,而IL-22则是一种稳态的稳态,仅是一种固定的稳态,仅是造成边界组织和组织的整体性启动性的,并且是造成边界组织和组织的构成的,并且是构成的,并且是构成的,并且是构成的,并且是构成的,并且是一种稳固的细胞影响。 [13,14]。IL-17和IL-22被认为是组织信号细胞因子,因为它们主要影响固体组织的细胞[12,15,16]。其受体的分布大大分歧。在人类中,IL-17的受体复合物在上皮,间质和造血细胞上广泛表达[13,17]。il-17和Th17细胞与各种器官(例如肝脏或皮肤)的促炎和纤维化条件有关[18,19]。

慢性乙型肝炎病毒感染孕妇的产后肝炎和宿主免疫
为了对胎儿产生免疫耐受,母体免疫系统与怀孕前相比会有一些变化。免疫耐受开始并发展于母体胎盘界面。在先天免疫中,蜕膜自然杀伤(dNK)细胞、巨噬细胞和树突状细胞在免疫耐受中起关键作用。在适应性免疫中,调节性T细胞(Treg)数量的适度增加和免疫抑制功能是免疫耐受所必需的。滋养层细胞和表达吲哚胺2,3-双加氧酶(IDO)的免疫细胞、表达HLA-G的滋养层细胞、Th1/Th2向Th2显性转变以及Th17/Treg向Treg显性转变有利于母胎免疫耐受。类固醇(雌激素和孕激素)和人绒毛膜促性腺激素(HCG)也通过诱导Treg细胞或上调免疫抑制细胞因子参与免疫耐受。慢性HBV感染者多数在妊娠前处于“HBV免疫耐受期”,妊娠期间肝脏疾病相对稳定。慢性HBV感染妇女分娩后,体内相对的免疫抑制状态发生逆转,Th1/Th2平衡中Th1占主导,Th17/Treg平衡中Th17占主导。分娩后,外周血中Treg数量减少,NK细胞数量增多且具有细胞毒性,肝脏NK细胞可能通过非抗原特异性机制引起肝脏炎症。分娩后,CD8+T细胞数量会回升,HBV特异性T细胞应答从妊娠期的功能障碍中恢复。在产后炎症的背景下,产后皮质醇的快速下降,特别是HBV DNA和细胞因子诱导的HBV特异性T细胞应答增强,是产后肝炎发生的主要原因。HBeAg阳性,特别是HBeAg<700 S/CO、HBV DNA>3-5Log 10 IU/ml是产后肝炎的危险因素。

CCL20/CCR6 趋化因子信号传导对于实验性自身免疫性脑脊髓炎小鼠多发性硬化症的发病机制并非至关重要
多发性硬化症是一种自身免疫性疾病,免疫系统会攻击神经髓鞘。致病性 Th17 细胞和调节性 Treg 细胞(均表达趋化因子受体 CCR6)之间的平衡对于确定疾病活动至关重要。据推测,由血脑屏障产生的 CCL20(CCR6 的同源配体)会将这些免疫细胞吸引到中枢神经系统 (CNS)。然而,CCR6 敲除 (KO) 小鼠中多发性硬化症实验模型的病理表型尚无定论,而 CCL20-KO 小鼠中尚未解决这一问题。为了解决这个问题,我们使用 CRISPR/Cas9 系统生成了 CCL20-KO 和 CCR6-KO 小鼠。与野生型 (WT) 小鼠相比,两种突变小鼠中实验性自身免疫性脑脊髓炎 (EAE) 慢性期的临床表型略有加重。 KO 小鼠和 WT 小鼠的中枢神经系统炎症细胞浸润和脱髓鞘相似。突变小鼠和 WT 小鼠的中枢神经系统 CD4 + T 细胞计数相同。突变小鼠和 WT 小鼠中枢神经系统 Th17 和 Treg 细胞的比例,或中枢神经系统 IL-17 和 TGF- b mRNA 表达没有显著差异。这些发现表明,CCL20/CCR6 介导的细胞迁移不一定是 EAE 发病所必需的,并且可能由其他趋化因子信号补偿。© 2022 Elsevier Inc. 保留所有权利。
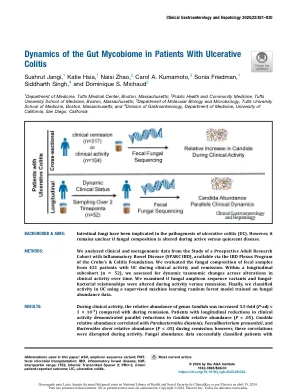
肠道结肠炎患者的肠道果酱组的动力学
ulcerative结肠炎(UC)是一种主要影响结肠的炎症性肠病(IBD)的一种形式,导致频繁复发,住院,手术和终生发病率提高。1 UC的病原体可能是由宿主微生物组稳态中的中断驱动的,特定细菌分类群的改变和与结肠渗透性相关的微生物产物的变化。2然而,针对肠道微生物组的疗法,包括益生菌,抗体或粪便微生物移植(FMT),在治疗UC方面具有适度且不一致的影响,因为在不完全表征的微生物组中存在个体间差异,因此在治疗UC方面具有适度的影响。3,4特别是,非细菌微生物王国在UC途径中的作用,尤其是真菌,以及操纵人群对疾病过程的影响。肠道真菌和真菌属念珠菌先前已与UC的发病机理有关。Dys-Biosis,其UC患者显示念珠菌中相对不断变化。5此外,包括Dectin-1在内的真菌抗原感应基因中的遗传多态性已与严重的UC形式有关。6小鼠中念珠菌的口服膨胀加剧了Th17介导的洪水,7和纤维形式的念珠菌可以激活炎症的肿瘤,并诱导结肠Th17的重音。8有趣的是,在移植前患有粪便念珠菌的FMT患者表现出对微生物疗法的有利反应。109我们先前还表明,在一小部分UC患者中,在内窥镜活性和缓解粪便中富含念珠菌(N¼53)。

细菌植物的优点和缺点
4。Ansaldo E,Slayden LC,Ching KL,Koch MA,Wolf NK,Plichta DR等。akkermansia粘膜粘膜在稳态期间诱导肠道适应性免疫反应。科学。2019; 364(6446):1179-1184。 5。 Sefik E,Geva-Zatorsky N,Oh S,Konnikova L,Zemmour D,McGuire AM等。 个体肠道共生体诱导RORγ +调节性T细胞的不同种群。 科学。 2015; 349(6251):993-997。 6。 Lathrop SK,Bloom SM,Rao SM,Nutsch K,Lio CW,Santacruz N等。 结肠共生微生物群对免疫系统的外围教育。 自然。 2011; 478(7368):250-254。 7。 Yang Y,Torchinsky MB,Gobert M,Xiong H,Xu M,Linehan JL等。 肠道Th17细胞对共生细菌抗原的聚焦特异性。 自然。 2014; 510(7503):152-156。 8。 Xu M,Pokrovskii M,Ding Y,Yi R,Au C,Harrison OJ等。 c- MAF依赖性调节性T细胞介导对肠道病原体的免疫耐受性。 自然。 2018; 554(7692):373-377。 9。 Chai JN,Peng Y,Rengarajan S,Solomon BD,AI TL,Shen Z等。 螺旋杆菌是体内平衡和炎症中结肠T细胞反应的有效驱动因素。 SCI免疫。 2017; 2(13):EAAL5068。 10。 Ivanov II,Atarashi K,Manel N,Brodie EL,Shima T,Karaoz U等。 通过分段丝状细菌诱导肠道Th17细胞。 单元格。 2009; 139(3):485-498。 11。2019; 364(6446):1179-1184。5。Sefik E,Geva-Zatorsky N,Oh S,Konnikova L,Zemmour D,McGuire AM等。个体肠道共生体诱导RORγ +调节性T细胞的不同种群。科学。2015; 349(6251):993-997。 6。 Lathrop SK,Bloom SM,Rao SM,Nutsch K,Lio CW,Santacruz N等。 结肠共生微生物群对免疫系统的外围教育。 自然。 2011; 478(7368):250-254。 7。 Yang Y,Torchinsky MB,Gobert M,Xiong H,Xu M,Linehan JL等。 肠道Th17细胞对共生细菌抗原的聚焦特异性。 自然。 2014; 510(7503):152-156。 8。 Xu M,Pokrovskii M,Ding Y,Yi R,Au C,Harrison OJ等。 c- MAF依赖性调节性T细胞介导对肠道病原体的免疫耐受性。 自然。 2018; 554(7692):373-377。 9。 Chai JN,Peng Y,Rengarajan S,Solomon BD,AI TL,Shen Z等。 螺旋杆菌是体内平衡和炎症中结肠T细胞反应的有效驱动因素。 SCI免疫。 2017; 2(13):EAAL5068。 10。 Ivanov II,Atarashi K,Manel N,Brodie EL,Shima T,Karaoz U等。 通过分段丝状细菌诱导肠道Th17细胞。 单元格。 2009; 139(3):485-498。 11。2015; 349(6251):993-997。6。Lathrop SK,Bloom SM,Rao SM,Nutsch K,Lio CW,Santacruz N等。结肠共生微生物群对免疫系统的外围教育。自然。2011; 478(7368):250-254。7。Yang Y,Torchinsky MB,Gobert M,Xiong H,Xu M,Linehan JL等。 肠道Th17细胞对共生细菌抗原的聚焦特异性。 自然。 2014; 510(7503):152-156。 8。 Xu M,Pokrovskii M,Ding Y,Yi R,Au C,Harrison OJ等。 c- MAF依赖性调节性T细胞介导对肠道病原体的免疫耐受性。 自然。 2018; 554(7692):373-377。 9。 Chai JN,Peng Y,Rengarajan S,Solomon BD,AI TL,Shen Z等。 螺旋杆菌是体内平衡和炎症中结肠T细胞反应的有效驱动因素。 SCI免疫。 2017; 2(13):EAAL5068。 10。 Ivanov II,Atarashi K,Manel N,Brodie EL,Shima T,Karaoz U等。 通过分段丝状细菌诱导肠道Th17细胞。 单元格。 2009; 139(3):485-498。 11。Yang Y,Torchinsky MB,Gobert M,Xiong H,Xu M,Linehan JL等。肠道Th17细胞对共生细菌抗原的聚焦特异性。自然。2014; 510(7503):152-156。 8。 Xu M,Pokrovskii M,Ding Y,Yi R,Au C,Harrison OJ等。 c- MAF依赖性调节性T细胞介导对肠道病原体的免疫耐受性。 自然。 2018; 554(7692):373-377。 9。 Chai JN,Peng Y,Rengarajan S,Solomon BD,AI TL,Shen Z等。 螺旋杆菌是体内平衡和炎症中结肠T细胞反应的有效驱动因素。 SCI免疫。 2017; 2(13):EAAL5068。 10。 Ivanov II,Atarashi K,Manel N,Brodie EL,Shima T,Karaoz U等。 通过分段丝状细菌诱导肠道Th17细胞。 单元格。 2009; 139(3):485-498。 11。2014; 510(7503):152-156。8。Xu M,Pokrovskii M,Ding Y,Yi R,Au C,Harrison OJ等。c- MAF依赖性调节性T细胞介导对肠道病原体的免疫耐受性。自然。2018; 554(7692):373-377。 9。 Chai JN,Peng Y,Rengarajan S,Solomon BD,AI TL,Shen Z等。 螺旋杆菌是体内平衡和炎症中结肠T细胞反应的有效驱动因素。 SCI免疫。 2017; 2(13):EAAL5068。 10。 Ivanov II,Atarashi K,Manel N,Brodie EL,Shima T,Karaoz U等。 通过分段丝状细菌诱导肠道Th17细胞。 单元格。 2009; 139(3):485-498。 11。2018; 554(7692):373-377。9。Chai JN,Peng Y,Rengarajan S,Solomon BD,AI TL,Shen Z等。螺旋杆菌是体内平衡和炎症中结肠T细胞反应的有效驱动因素。SCI免疫。 2017; 2(13):EAAL5068。 10。 Ivanov II,Atarashi K,Manel N,Brodie EL,Shima T,Karaoz U等。 通过分段丝状细菌诱导肠道Th17细胞。 单元格。 2009; 139(3):485-498。 11。SCI免疫。2017; 2(13):EAAL5068。 10。 Ivanov II,Atarashi K,Manel N,Brodie EL,Shima T,Karaoz U等。 通过分段丝状细菌诱导肠道Th17细胞。 单元格。 2009; 139(3):485-498。 11。2017; 2(13):EAAL5068。10。Ivanov II,Atarashi K,Manel N,Brodie EL,Shima T,Karaoz U等。 通过分段丝状细菌诱导肠道Th17细胞。 单元格。 2009; 139(3):485-498。 11。Ivanov II,Atarashi K,Manel N,Brodie EL,Shima T,Karaoz U等。通过分段丝状细菌诱导肠道Th17细胞。单元格。2009; 139(3):485-498。 11。2009; 139(3):485-498。11。Bilate AM,Bousbaine D,Mesin L,Agudelo M,Leube J,Kratzert A等。来自克隆T细胞前体的调节和上皮内T细胞的组织特异性出现。SCI免疫。 2016; 1(2):EAAF7471。 12。 Bilate Am,Lafaille JJ。 在免疫耐受性中诱导的CD4+ FOXP3+调节T细胞。 Annu Rev Immunol。 2012; 30:733-758。 13。 页岩M,Schiering C,Powrie F.肠炎中的CD 4+ T细胞子集。 Immunol Rev. 2013; 252(1):164-182。 14。 Sujino T,伦敦M,Hoytema van Konijnenburg DP,Rendon T,Buch T,Silva HM等。 调节和上皮内CD4+ T细胞的组织适应控制肠道炎症。 科学。 2016; 352(6293):1581-1586。 15。 Reis BS,Rogoz A,Costa-Pinto FA,Taniuchi I,MucidaD。转录因子Runx3和ThPOK的相互表达调节肠道CD4+ T细胞免疫。 nat免疫。 2013; 14(3):271-280。SCI免疫。2016; 1(2):EAAF7471。 12。 Bilate Am,Lafaille JJ。 在免疫耐受性中诱导的CD4+ FOXP3+调节T细胞。 Annu Rev Immunol。 2012; 30:733-758。 13。 页岩M,Schiering C,Powrie F.肠炎中的CD 4+ T细胞子集。 Immunol Rev. 2013; 252(1):164-182。 14。 Sujino T,伦敦M,Hoytema van Konijnenburg DP,Rendon T,Buch T,Silva HM等。 调节和上皮内CD4+ T细胞的组织适应控制肠道炎症。 科学。 2016; 352(6293):1581-1586。 15。 Reis BS,Rogoz A,Costa-Pinto FA,Taniuchi I,MucidaD。转录因子Runx3和ThPOK的相互表达调节肠道CD4+ T细胞免疫。 nat免疫。 2013; 14(3):271-280。2016; 1(2):EAAF7471。12。Bilate Am,Lafaille JJ。在免疫耐受性中诱导的CD4+ FOXP3+调节T细胞。Annu Rev Immunol。 2012; 30:733-758。 13。 页岩M,Schiering C,Powrie F.肠炎中的CD 4+ T细胞子集。 Immunol Rev. 2013; 252(1):164-182。 14。 Sujino T,伦敦M,Hoytema van Konijnenburg DP,Rendon T,Buch T,Silva HM等。 调节和上皮内CD4+ T细胞的组织适应控制肠道炎症。 科学。 2016; 352(6293):1581-1586。 15。 Reis BS,Rogoz A,Costa-Pinto FA,Taniuchi I,MucidaD。转录因子Runx3和ThPOK的相互表达调节肠道CD4+ T细胞免疫。 nat免疫。 2013; 14(3):271-280。Annu Rev Immunol。2012; 30:733-758。 13。 页岩M,Schiering C,Powrie F.肠炎中的CD 4+ T细胞子集。 Immunol Rev. 2013; 252(1):164-182。 14。 Sujino T,伦敦M,Hoytema van Konijnenburg DP,Rendon T,Buch T,Silva HM等。 调节和上皮内CD4+ T细胞的组织适应控制肠道炎症。 科学。 2016; 352(6293):1581-1586。 15。 Reis BS,Rogoz A,Costa-Pinto FA,Taniuchi I,MucidaD。转录因子Runx3和ThPOK的相互表达调节肠道CD4+ T细胞免疫。 nat免疫。 2013; 14(3):271-280。2012; 30:733-758。13。页岩M,Schiering C,Powrie F.肠炎中的CD 4+ T细胞子集。Immunol Rev.2013; 252(1):164-182。 14。 Sujino T,伦敦M,Hoytema van Konijnenburg DP,Rendon T,Buch T,Silva HM等。 调节和上皮内CD4+ T细胞的组织适应控制肠道炎症。 科学。 2016; 352(6293):1581-1586。 15。 Reis BS,Rogoz A,Costa-Pinto FA,Taniuchi I,MucidaD。转录因子Runx3和ThPOK的相互表达调节肠道CD4+ T细胞免疫。 nat免疫。 2013; 14(3):271-280。2013; 252(1):164-182。14。Sujino T,伦敦M,Hoytema van Konijnenburg DP,Rendon T,Buch T,Silva HM等。调节和上皮内CD4+ T细胞的组织适应控制肠道炎症。科学。2016; 352(6293):1581-1586。 15。 Reis BS,Rogoz A,Costa-Pinto FA,Taniuchi I,MucidaD。转录因子Runx3和ThPOK的相互表达调节肠道CD4+ T细胞免疫。 nat免疫。 2013; 14(3):271-280。2016; 352(6293):1581-1586。15。Reis BS,Rogoz A,Costa-Pinto FA,Taniuchi I,MucidaD。转录因子Runx3和ThPOK的相互表达调节肠道CD4+ T细胞免疫。nat免疫。2013; 14(3):271-280。2013; 14(3):271-280。

肠道衍生的醋酸酯促进具有抗炎作用的B10细胞
引言使自身免疫性疾病(如类风湿关节炎)可能是由于免疫耐受性的分解而引起的。这种崩溃的原因尚不完全清楚,但关键特征是proinflam-sy-sy-procatory Th17和Th1 T细胞子集的扩展,抗炎Treg子集的伴随降低(1)。B细胞的一个小子集,该子集释放抗炎细胞因子IL-10,在免疫耐受性中也起着重要作用。这些调节B细胞(也称为B10细胞)促进了Treg分化,同时抑制Th1和Th17细胞(2)。在临床前和临床研究中,B10细胞在耐受性中的关键作用已确定。的确,小鼠中缺乏B10细胞会加剧关节炎的发展(3),而在实验性自身免疫性脑炎的小鼠模型中,B10细胞的继发性转移显着降低了自身免疫性疾病的严重程度(4),结肠炎(5)和关节炎(6)。类风湿关节炎患者的特征是B10细胞的比例显着降低,其耐受功能受损(7)。这些患者中B10细胞的比例也与疾病活性成反比(7)。可以修复这些患者中B10细胞的功能和比例,因此可以用作一种可能适用于其他炎症性疾病的新治疗方法。在小鼠中,B10细胞主要来自腹膜祖细胞B1A细胞。 b10细胞在脾脏中的较小程度较小,源自B2祖细胞B细胞,尤其是T2-边缘区的前体(8)。在小鼠中,B10细胞主要来自腹膜祖细胞B1A细胞。b10细胞在脾脏中的较小程度较小,源自B2祖细胞B细胞,尤其是T2-边缘区的前体(8)。在人类中,血液来源的B10细胞主要衍生自CD24 HI CD38 HI和CD24 HI CD27 + B细胞(9)。B10细胞分化是由环境线索驱动的,促炎细胞因子(例如IL-21)具有

维生素C
关于Covid-19,据信免疫反应涉及炎性和抗炎细胞因子之间的复杂相互作用。 几项研究报道了由COVID-19的患者的细胞因子和趋化因子的异常水平(8)。 大流行,Blanco-Mello等。 表明,合并症患者的不适当和弱免疫反应出现的频率更高(9)。 当人体对病毒的自然防御措施被削弱并大量产生炎症性细胞因子时,SARS-COV-2感染可能会有一个转折点(9)。 在许多患有严重兴奋的促炎细胞因子(例如肿瘤坏死因子(TNF))的血清水平较高的患者中,与患有轻度疾病(10、11)相比,已经观察到较高的肿瘤坏死因子(TNF) - α,α,白介素(IL)-1β,IL-2和IL-6以及IL-8等趋化因子。 这表明COVID-19的发病机理涉及T-Helper细胞(Th)亚群(包括TH1,TH2和TH17)以及调节性T细胞(Tregs)的不平衡。关于Covid-19,据信免疫反应涉及炎性和抗炎细胞因子之间的复杂相互作用。几项研究报道了由COVID-19的患者的细胞因子和趋化因子的异常水平(8)。大流行,Blanco-Mello等。表明,合并症患者的不适当和弱免疫反应出现的频率更高(9)。当人体对病毒的自然防御措施被削弱并大量产生炎症性细胞因子时,SARS-COV-2感染可能会有一个转折点(9)。在许多患有严重兴奋的促炎细胞因子(例如肿瘤坏死因子(TNF))的血清水平较高的患者中,与患有轻度疾病(10、11)相比,已经观察到较高的肿瘤坏死因子(TNF) - α,α,白介素(IL)-1β,IL-2和IL-6以及IL-8等趋化因子。这表明COVID-19的发病机理涉及T-Helper细胞(Th)亚群(包括TH1,TH2和TH17)以及调节性T细胞(Tregs)的不平衡。

关于EFSA和BFR在EFSA Bisphenol A(BPA)意见之间发散意见的报告
存在。既没有得出终点的研究(Luo等,2016),也没有其他长期暴露研究(参见例如tyl等,2008,Delclos等,2014年,在Clarity Project中进行的研究或流行病学研究报告了诸如炎症之类的不良根尖作用。对于此终点,不存在任何认可的不良结果途径。因此,BFR认为,新型的中间端点“脾脏中的Th17细胞百分比增加”似乎没有足够的合理性和确定为动物或人类不良健康结果的预测指标,并且认为它不适合衍生HBGV。沿着这些行,BFR指出,此端点的选择与EFSA使用的WHO/IPCS定义(WHO/IPCS,2009年)一致。bfr认为,动物模型中的中间端点“脾脏Th17细胞百分比增加”目前尚未足够合理地作为动物和人类不良健康结果的预测指标。因此,BFR认为选择该中间端点是一种范式转移:它导致考虑与某种物质暴露有关的人类健康风险的证据,以考虑可能的逆境,这可能最终在体内表现出来。bfr认为这与一般实践不符。此外,根据BFR,在风险评估过程的每个步骤中都使用了保守的最坏情况假设(例如bfr不同意EFSA进行的危害表征,因此不支持TDI和以下风险表征。选择关注的效果,整个证据的重量(祸)过程,毒理因子的选择以及量化剩余不确定性的方法,导致过度保守的HBGV。efsa强调,所使用的逆境的定义(WHO/IPCS,2009年)没有提及,也没有要求对效果的根尖性被视为不利。在设置HBGV时,EFSA考虑了与可能的根尖不良影响有关系的影响,因此可能在毒理学上相关。但是,这种影响不一定需要与一对一因果关系中的顶端终点相关。efsa包括使用中间端点,认为与不良结果具有明显的因果关系(AO)。efsa认为,通过根据应用方案加权总体证据体,可以确定中间效应与不利结果之间的联系,即使AO不一定在所考虑的研究的设计和范围内表达,并且不一定在单个(指南)研究中得到证实。意见中审查的证据(第3.1.3节)以及对BPA效应的越来越多的科学证据(请参阅对附件N中评论30的回答)清楚地表明,Th17细胞百分比的增量及其全白蛋白的增量表明,与免疫系统一致的疾病相关的动物均与抗症状相关的疾病的差异性增强,并涉及各种免疫力的疾病,并涉及无效的疾病,并涉及侵蚀性,并涉及无效的疾病。人类(例如牛皮癣,糖尿病,多发性硬化症,嗜中性哮喘等)。这种证据也存在于几种动物中