XiaoMi-AI文件搜索系统
World File Search SystemTTTV
基因组编辑-Alt -R为CAS12A(CPF1)超核酸酶-Net
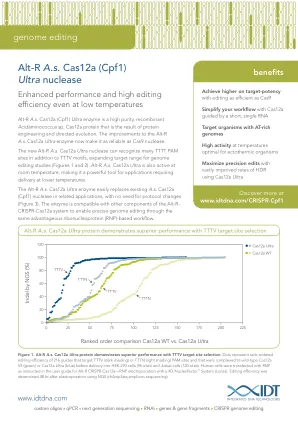
图1。Alt-R A.S. CAS12A超蛋白在TTTV目标位点选择方面表现出卓越的性能。 DOTS代表了216个指南的等级编辑效率,该指南靶向TTTV(深色阴影)或TTTN(浅色阴影)PAM站点,并且在输送到HEK-293细胞(96个位点)和陪同下(96个位点)和陪同细胞(96个位点)和野生型CAS12A V3(绿色)V3(绿色)或Cas12a Ultra(蓝色)。 如Alt-R CRISPR-CAS12A用户指南中所指示的RNP 人类细胞使用4D-Nucleofector™System(LONZA)的RNP电穿孔。 使用NGS(Rhampseq Amplicon测序)进行电穿孔后48小时编辑效率。Alt-R A.S. CAS12A超蛋白在TTTV目标位点选择方面表现出卓越的性能。DOTS代表了216个指南的等级编辑效率,该指南靶向TTTV(深色阴影)或TTTN(浅色阴影)PAM站点,并且在输送到HEK-293细胞(96个位点)和陪同下(96个位点)和陪同细胞(96个位点)和野生型CAS12A V3(绿色)V3(绿色)或Cas12a Ultra(蓝色)。人类细胞使用4D-Nucleofector™System(LONZA)的RNP电穿孔。使用NGS(Rhampseq Amplicon测序)进行电穿孔后48小时编辑效率。编辑效率。

改进的 LbCas12a 变体具有改变的 PAM 特异性......
Cas12a(以前称为 Cpf1)核酸酶在基因组工程中的广泛使用受到它们需要相当长的 TTTV 原型间隔区相邻基序 (PAM) 序列的限制。在这里,我们旨在放宽这些 PAM 限制,并通过将其相应的 RR 和 RVR 变体的突变与改变的 PAM 特异性相结合,生成了在哺乳动物和植物细胞中活跃的四种 Cas12a 直系同源物的新型 PAM 突变变体。选择表现出最高活性的 LbCas12a-RVRR,使用基于质粒的测定法深入表征其在哺乳动物细胞中的 PAM 偏好。LbCas12a-RVRR 的共识 PAM 序列类似于 TNTN 基序,但也包括 TACV、TTCV CTCV 和 CCCV。经发现,改良的 LbCas12a (impLbCas12a) 中的 D156R 突变以 PAM 依赖的方式进一步提高了该变体的活性。由于 impLbCas12a 和最近报道的 enAsCas12a 变体的 PAM 偏好重叠但仍有差异,它们相互补充,为基因组编辑和转录组调节应用提供了更高的效率。

改进的 LbCas12a 变体具有改变的 PAM 特异性......
Cas12a(以前称为 Cpf1)核酸酶在基因组工程中的广泛使用受到它们需要相当长的 TTTV 原型间隔区相邻基序 (PAM) 序列的限制。在这里,我们旨在放宽这些 PAM 限制,并通过将其相应的 RR 和 RVR 变体的突变与改变的 PAM 特异性相结合,生成了在哺乳动物和植物细胞中活跃的四种 Cas12a 直系同源物的新型 PAM 突变变体。选择表现出最高活性的 LbCas12a-RVRR,使用基于质粒的测定法深入表征其在哺乳动物细胞中的 PAM 偏好。LbCas12a-RVRR 的共识 PAM 序列类似于 TNTN 基序,但也包括 TACV、TTCV CTCV 和 CCCV。经发现,改良的 LbCas12a (impLbCas12a) 中的 D156R 突变以 PAM 依赖的方式进一步提高了该变体的活性。由于 impLbCas12a 和最近报道的 enAsCas12a 变体的 PAM 偏好重叠但仍有差异,它们相互补充,为基因组编辑和转录组调节应用提供了更高的效率。

CRISPR/CAS12A基因组编辑工具箱...
甲烷古细菌在全球碳循环中起着重要作用,可以作为CO 2和其他一碳基质的燃料和化学物质生物技术生产的宿主生物。甲藻菌的乙酰硫酸酯因其较大的基因组,多功能底物范围和可用的遗传工具而被广泛研究为甲烷原模型。也已经证明了通过CRISPR/CAS9在M. acetivorans中进行基因组编辑。在这里,我们描述了一个用户友好的CRISPR/CAS12A工具箱,该工具箱识别富含T的PAM序列。该工具箱可以管理3,500 bp(即淘汰整个Frhadgb操纵子)和异源基因插入的缺失,正率超过80%。cas12a介导的多重基因组编辑用于在一轮编辑中编辑染色体上的两个单独的位点。达到了100 bp的双重删除,正确编辑了8/8的转化子。在一个位置同时删除100 bp,并用2,400 bp的UIDA表达盒在另一个位置替换100 bp,可在单独的位置上获得5/6个正确编辑的转换物。我们的CRISPR/CAS12A工具箱可实现可靠的基因组编辑,并且可以与先前报道的基于CAS9的基于CAS9的系统并行使用,用于甲状腺素物种的基因工程。

Cas12a 直系同源物和工程变体的高通量分析,以增强基因组编辑活性
摘要:CRISPR/Cas12a(以前称为 Cpf1)是一种 RNA 引导的 VA 类 CRISPR 系统内切酶,为基因组工程提供了一种有前途的工具。目前已鉴定出 10 多个 Cas12a 直系同源物,并用于人类细胞的基因编辑。然而,新兴 Cas12a 直系同源物之间的功能多样性仍未得到充分探索。本文,我们通过构建包含 40,000 多个引导 RNA 的基因组整合、自切割、配对 crRNA 靶标文库,报告了 16 个 Cas12a 直系同源物在人类细胞中的编辑活性的高通量比较分析。三种 Cas12a 候选物由于其结构紧凑且编辑效率与 AsCas12a 和 LbCas12a 相当而表现出良好的潜力,而 AsCas12a 和 LbCas12a 的特征已得到充分表征。我们通过结构引导的蛋白质工程生成了三种精氨酸替代变体 (3Rv):BsCas12a-3Rv (K155R/N512R/K518R)、PrCas12a-3Rv (E162R/N519R/K525R) 和 Mb3Cas12a-3Rv (D180R/N581R/K587R)。与野生型 Cas12a 效应子相比,这三种 Cas12a 变体均表现出增强的编辑活性和扩大的靶向范围 (NTTV、NTCV 和 TRTV)。三种 Cas12a 变体之间的碱基偏好分析表明,PrCas12a-3Rv 在具有典型 PAM TTTV 和非典型 PAM TTCV 的靶位点上表现出最高活性,而 Mb3Cas12a-3Rv 表现出与其他变体不同的识别特征,在 PAM TATV 的 -3 位置和 PAM ATCV 的 -4 位置容纳更多的核苷酸 A。因此,扩展的 Cas12a 工具箱和对 Cas12a 活动的更好理解应该有助于它们在基因组工程中的应用。

一种用于体内基因组编辑的新型紧凑型 Cas12a 变体
为了使通过腺相关病毒 (AAV) 载体进行的基因治疗取得成功,载体基因组大小是一个主要制约因素,因为它会阻止将较大的转基因包装到二十面体病毒衣壳中。在基于 CRISPR 的基因组编辑背景下,已经描述了多种紧凑型 Cas 变体,例如 Cas12j (CasPhi) 或 Cas12f (CasMINI) [ 1 , 2 ],它们可用于通过 AAV 递送进行体内基因组编辑。在 Wang 等人的研究中 [ 3 ],一种来自丹毒菌 (EbCas12a) 的新型紧凑型 Cas12a 变体被表征为当通过点突变 (enEbCas12a) 改进时,显示出与其他 Cas12a 变体相似的编辑效率 [ 3 ]。值得注意的是,EbCas12a 的编码序列比下一个更大的已表征 Cas12a 变体小约 150 bp(图 1),有利于将其容纳在“一体化” AAV 载体中,该载体在单个 AAV 模板上提供 Cas12a 和 crRNA。通过使用单一载体给药,作者证明了新型 AAV-enEbCas12a 载体介导体内基因组编辑的能力,从而为不断扩展的 CRISPR 工具箱增加了一个新条目。首先,作者通过体外切割试验表征了 EbCas12a,并确定 TTTV(V = G、C 或 A)为 PAM 序列,这与其他已报道的 Cas12a 系统类似。接下来,EbCas12 被证明在培养的哺乳动物细胞中具有功能性,这通过两个报告基因和各种基因组位点的切割得到证实。然而,与其他 Cas12a 变体(例如常用的 AsCas12a 和 LbCas12a)相比,EbCas12a 效率较低。因此,为了放宽 PAM 序列限制并提高 AsCas12a 的编辑效率,作者们在 EbCas12a 中替换了一个氨基酸,以建立 PAM 近端 DNA 接触,从而产生了变体 enEbCas12a。事实上,这种单点突变将基因组位点的编辑效率提高了约 2 倍。然而,与此同时,放宽 PAM 序列限制可能会增加 enEbCas12a 脱靶编辑的风险。为了通过实验检验这一担忧,对脱靶编辑事件进行了全基因组分析。值得注意的是,检测到的脱靶位点数量为

CRISPR 筛选:研究病毒的分子工具——...
病毒是专性病原体,利用宿主细胞机制进行复制。宿主细胞可以识别病毒并激活抗病毒反应。揭示影响病毒感染的因素有助于发现新的候选药物。使用有助于抗病毒免疫反应的特异性免疫激动剂是治疗感染的另一种方法。最近,使用新的综合分子工具(例如成簇的规律间隔的短回文重复序列 (CRISPR) 筛选)研究宿主细胞相互作用并确定开发新抗病毒药物的关键靶标已成为可能。在过去十年中,CRISPR/CRISPR 相关蛋白 (Cas) 系统已被用于基因组编辑。仅当基因组靶标后面跟着一个原间隔区相邻基序 (PAM) 序列(Cas9 蛋白为 NGG,Cas12a 蛋白为 TTTV)时,Cas 蛋白才会使用 II 型 CRISPR 系统中的单向导 RNA (sgRNA) 和 V 型系统中的 CRISPR RNA (crRNA)(为简单起见,在本综述中称为 gRNA)识别靶位点。在识别靶位点后,Cas 蛋白会解开 DNA 链,形成 R 环结构,并切断两条链,导致 DNA 双链断裂 (DSB)。利用位点特异性诱变,已经生成了具有核酸酶钝结构域的 Cas 内切酶变体,称为核酸酶失活 Cas (dCas) 蛋白。dCas 保留了结合目标位点的能力,但不能将 DSB 引入 DNA。将不同的功能域融合到 dCas 蛋白上,可将其转化为具有多种功能的分子“瑞士军刀”,例如单核苷酸编辑和调节转录和表观遗传学 [1]。通过不同的 CRISPR/Cas 系统激活或抑制靶基因转录已被广泛用于破坏单个基因和研究病毒-宿主相互作用 [2]。通过设计和合成数千个针对多个目标基因或基因组中所有基因的 gRNA,可以使用 CRISPR/Cas

使用新型 Lb Cas12a 变体对大麦进行高效基因组编辑以及 sgRNA 结构的影响 Tom Lawrenson、Alison Hinchliffe、Macarena
CRISPR、Cas12a、CPF1、大麦、诱变、单子叶植物、基因组编辑摘要我们报告了首次成功、高效使用大麦中的 Lb Cas12a,并描述了两种新型 Cas12a 变体的开发和应用。总共我们使用二十种不同的指南比较了五种编码序列 (CDS) 变体,包括两种新型变体和两种指南架构,针对 5 种不同的靶基因。我们发现不同 CDS 版本 (0-87%) 和指南架构 (0-70%) 之间的编辑效率存在很大差异,并且表明我们的两个新型 CDS 版本在该物种的测试中大大优于其他版本。我们展示了产生的突变的遗传性。我们的研究结果强调了优化单个物种的 CRISPR 系统的重要性,并可能有助于在其他单子叶植物中使用 Lb Cas12a。正文 毛螺菌科细菌 Cas12a (Lb Cas12a) 可能是继化脓性链球菌 Cas9 (Sp Cas9) 之后植物基因组编辑中第二广泛使用的可编程核酸酶,并且具有一些潜在优势。首先,由于其对 TTTV PAM 的要求与 NGG 的 Sp Cas9 要求不同,它可用于 GC 沙漠,而 GC 沙漠通常存在于内含子、UTR 和启动子区域中。其次,Lb Cas12a 通常比 Sp Cas9 产生更大的缺失,这可能在缺失研究中有用。第三,虽然 Sp Cas9 在靶标的 PAM 近端切割产生平端,但 Lb Cas12a 在 PAM 远端区域切割产生粘端;这两个特征可能解释了使用 Lb Cas12a 实现的基因靶向发生率更高 (Wolter 和 Puchta,2019)。已知在植物中起作用的三种版本的 Lb Cas12a 针对一个大麦靶标进行了测试。首先,是水稻优化的编码序列 (CDS) (Os Cas12a) (Tang et al., 2017);其次是人类优化的 CDS (Hs Cas12a),在双子叶植物中具有功能 (Bernabé-Orts et al., 2019);第三是拟南芥优化的 CDS,包含 D156R“耐高温”突变 (tt At Cas12a) (Schindele and Puchta, 2020)。我们还创建了两个新版本,携带 D156R 突变的 Hs Cas12a (tt Hs Cas12a) 和携带 8 个内含子的 tt At Cas12 (tt At Cas12+int)。这些内含子之前曾显著提高过 Sp Cas9 的效率(Grutzner 2021),因此我们使用相同的在线工具(NetGene2 - 2.42 - Services - DTU Health Tech)在我们的 tt At Cas12+int 设计中为拟南芥选项获得了较高的剪接置信度。