XiaoMi-AI文件搜索系统
World File Search SystemTadA

利用 TadA 的下一代胞嘧啶碱基编辑器
• 脱氨酶的定向进化 • PAM 变体碱基编辑器 • 定向进化 Cas9 以创建用于 BE 的非 NGG PAM 变体 • 密码子、NLS 和接头优化 • 环状置换体和镶嵌碱基编辑器 • DNA 脱靶评估 • RNA 脱靶评估 • 旁观者编辑最小化 • 引导 RNA 工程 • 离体和体内 BE 递送 • 最小化脱靶活性的工程 BE • HSC、肝细胞和 T 细胞的离体碱基编辑 • ABE 的低温电子显微镜结构 • 小鼠体内碱基编辑 • 非人类灵长类动物体内编辑

由 TadA 变体生成的改进的胞嘧啶碱基编辑器
胞嘧啶碱基编辑器 (CBE) 可实现可编程的基因组 C·G 到 T·A 转换突变,通常包含经过修饰的 CRISPR-Cas 酶、天然存在的胞嘧啶脱氨酶和尿嘧啶修复抑制剂。先前的研究表明,利用天然存在的胞嘧啶脱氨酶的 CBE 可能导致无引导的全基因组胞嘧啶脱氨。尽管随后报道了可减少随机全基因组脱靶的改进型 CBE,但这些编辑器的靶向性能可能不理想。本文,我们报告了使用 TadA 的工程变体 (CBE-T) 的 CBE 的生成和表征,这些变体可在序列多样的基因组位点上实现高靶向 C·G 到 T·A,在原代细胞中表现出强大的活性,并且不会导致全基因组突变的可检测升高。此外,我们报道了胞嘧啶和腺嘌呤碱基编辑器 (CABE),它们可催化 A 到 I 和 C 到 U 编辑 (CABE-T)。与 ABE 一起,CBE-T 和 CABE-T 可使用实验室进化的 TadA 变体对所有转换突变进行可编程安装,与之前报道的 CBE 相比,这些变体具有更好的特性。

Tada 伐木服务
(7) 其他 A、双方当事人在合同上签字盖章后,合同成立。 B.确定中标时,按投标文件中记载的金额加上相当于该金额10%的金额(该金额中存在不足1日元的小数部分,小数部分向下舍入)为。考虑到中标价格,因此投标人,无论是应税企业还是免税企业,都需要支付相当于预计合同金额的110/110的金额。金额应在投标文件中注明。 C、参加投标者须提交资格审查结果通知书复印件。 D、如需委托代理人参加投标,请提交授权委托书。 如果您希望参加投标,请在指定地点查看《投标与合同指南》,并填写投标文件中的必要信息。 (f) 通过邮寄方式接受投标。此时,在信封上写清楚“投标主题”,附上资格审查结果通知书复印件,并在周三17:00之前提交陆上自卫队上富良野警备队第344会计中队的合同, 2020年11月6日,请确保已送达团体,如遇此情况,请电话联系负责人确认抵达。 G. 邮寄投标的重新投标将另行注明。 在投标表格底部的空白处,有一条消息:“我们的公司(如果您是私人或个人)或我们的组织(如果您是组织)已同意条款和条件根据上述公告,我会投标。我们还承诺按照《招标和合同准则》的规定消除有组织犯罪。 ”。 投标、标准等咨询请联系陆上自卫队上富良野警备队第 344 会计中队合同组(负责人:石崎) 电话:0167-45-3101(内线:2343) 传真:0167-45-3101(分机:2660) ) (八) 公示地点和期限发布地点:旭川县、上富良野驻防会计队、旭川商工会议所、上富良野商工会议所北部地区会计队主页 http://www.mod.go.jp/gsdf.nae/fin/ (a) 发布期间:Rei 2020年10月22日星期二 - 2020年11月7日星期四

这是以下文章的同行评审版本:Ranzau、Brodie L.、Rallapalli、Kartik L.、Evanoff、Mallory、Paesani、Francesco、Komor、Ale
碱基编辑器是一种基因组编辑工具,可通过对 DNA 中的核碱基进行化学修饰来实现位点特异性碱基转换。腺嘌呤碱基编辑器 (ABE) 利用腺苷脱氨酶将目标腺苷修饰为肌苷中间体,从而将 DNA 中的 A•T 转换为 G•C 碱基对。由于缺乏可以修饰 DNA 的天然腺苷脱氨酶,ABE 是从 tRNA 脱氨酶 TadA 进化而来的。之前利用由野生型 (wt) TadA 组成的 ABE 进行的实验未显示对 DNA 的可检测活性,因此需要定向进化以使该酶能够接受 DNA 作为底物。在这里,我们表明 wtTadA 可以在细菌和哺乳动物细胞中的 DNA 中进行碱基编辑,对 TAC 的序列基序有严格的要求。我们利用这一发现优化了报告基因检测,以检测低至 0.01% 的碱基编辑水平。最后,我们将该分析与完整 ABE:DNA 复合物的分子动力学模拟结合使用,以更好地了解突变 TadA 变体的序列识别如何随着它们积累突变而变化,从而更好地编辑 DNA 底物。

加州大学圣地亚哥分校
碱基编辑器是一种基因组编辑工具,可通过对 DNA 中的核碱基进行化学修饰来实现位点特异性碱基转换。腺嘌呤碱基编辑器 (ABE) 利用腺苷脱氨酶将目标腺苷修饰为肌苷中间体,从而将 DNA 中的 A•T 转换为 G•C 碱基对。由于缺乏可以修饰 DNA 的天然腺苷脱氨酶,ABE 是从 tRNA 脱氨酶 TadA 进化而来的。之前利用由野生型 (wt) TadA 组成的 ABE 进行的实验未显示对 DNA 的可检测活性,因此需要定向进化以使该酶能够接受 DNA 作为底物。在这里,我们表明 wtTadA 可以在细菌和哺乳动物细胞中的 DNA 中进行碱基编辑,对 TAC 的序列基序有严格的要求。我们利用这一发现优化了报告基因检测,以检测低至 0.01% 的碱基编辑水平。最后,我们将该分析与完整 ABE:DNA 复合物的分子动力学模拟结合使用,以更好地了解突变 TadA 变体的序列识别如何随着它们积累突变而变化,从而更好地编辑 DNA 底物。


DNA测序方法 div>
添加尿素,并在70°C下进行电泳,以便消除所有氢连接。 div>这些条件可确保仅根据DNA的大小和序列将DNA片段分开,然后可以使用凝胶功能图直接读取•可以将300分开,最多连续400个连续 div div>

大规模基因组和转录组测序分析揭示了植物中高活性腺嘌呤碱基编辑器诱导的突变景观
摘要背景:利用最近开发的 tRNA 腺苷脱氨酶 (TadA8e 和 TadA9) 改造的高活性腺嘌呤碱基编辑器 (ABE) 表现出强大的碱基编辑活性,但引发了人们对脱靶效应的担忧。结果:在本研究中,我们对 ABE8e 和 ABE9 诱导的水稻 DNA 和 RNA 突变进行了全面评估。对用四种 ABE(包括 SpCas9n-TadA8e、SpCas9n-TadA9、SpCas9n-NG-TadA8e 和 SpCas9n-NG-TadA9)转化的植物进行全基因组测序分析表明,含有 TadA9 的 ABE 导致更多数量的脱靶 A 到 G (A>G) 单核苷酸变体 (SNV),而含有 CRISPR/SpCas9n-NG 的 ABE 导致水稻基因组中脱靶 SNV 总数更高。对携带 ABE 的 T-DNA 的分析表明,在 T-DNA 整合到植物基因组之前和/或之后可以引入靶向突变,在 ABE 整合到基因组之后会形成更多的脱靶 A>G SNV。此外,我们在 ABE 表达高的植物中检测到脱靶 A>G RNA 突变,但在 ABE 表达低的植物中未检测到。脱靶 A>G RNA 突变倾向于聚集,而脱靶 A>G DNA 突变很少聚集。结论:我们的研究结果表明 Cas 蛋白、TadA 变体、ABE 的时间表达和 ABE 的表达水平对水稻中的 ABE 特异性有影响,这为了解 ABE 的特异性提供了见解,并提出了除改造 TadA 变体之外增加 ABE 特异性的其他方法。

血脑屏障的生物学和模型在CRISPR中完善定位
CRISPR (clustered regularly interspaced short palindromic repeats) is a natural bacterial defense system against bacteriophage infection that has recently been harnessed for genome and tran- scriptome editing in a wide range of organisms based on the generation of double-strand DNA breaks (DSBs) and RNA cleavage (3, 24, 32, 47, 52, 58, 73, 76, 79, 91,127)。是根据工程II(CAS9)和VI型(CAS13)可编程核酸酶,DNA和RNA基础编辑,质量编辑以及CRISPR干扰/激活(CRISPRI/A)编辑(CRISPRI/A)编辑(CRISPRI/A)编辑,启用与基本疾病的校正和安装基本疾病的校正和安装,40个基本疾病的突变(30; 69–71、87、105、115、135),例如转录扰动(138)和表观遗传调节(94)。这些基于DNA的编辑器是通过没有DSB活性的死亡CAS9(DCAS9)或CAS9 Nickase(CAS9N)的融合而生成的,只有对胞嘧啶脱氨酶的活性(例如,APOBEC和C-TO-T编辑的APOBEC和辅助)或trans-FER RNA(TRNA)腺苷(TRNA)腺苷氨基氨基酶(例如,tada)(例如,tada)(37)(37)(37)(37)。RNA编辑系统是通过将DCAS13B/DCAS13D/DCAS13X融合而成的,没有RNA裂解活性与腺苷脱氨酶结构域(例如,ADAR2 DD用于A-TO-I编辑)或工程型胞质Deam-Inase Inase Insaine(例如,ADAR2DD)的87,C-TON 7,c-us-n.7,c.-ty 7,c c. 47,c. 47,c.-ty 7,c-ty 7,c-ty 7,c-us-c.-edy in 13,c-u-u--u-u-udy in 13,c-u-udy in 34,c-u-u--为了启用序列特异性基因组调节,DCAS蛋白还融合到多个基因调节效应子,例如逆转录酶(10),转录阻遏物和激活剂(40,101)和表观遗传性调节器(17,99)。
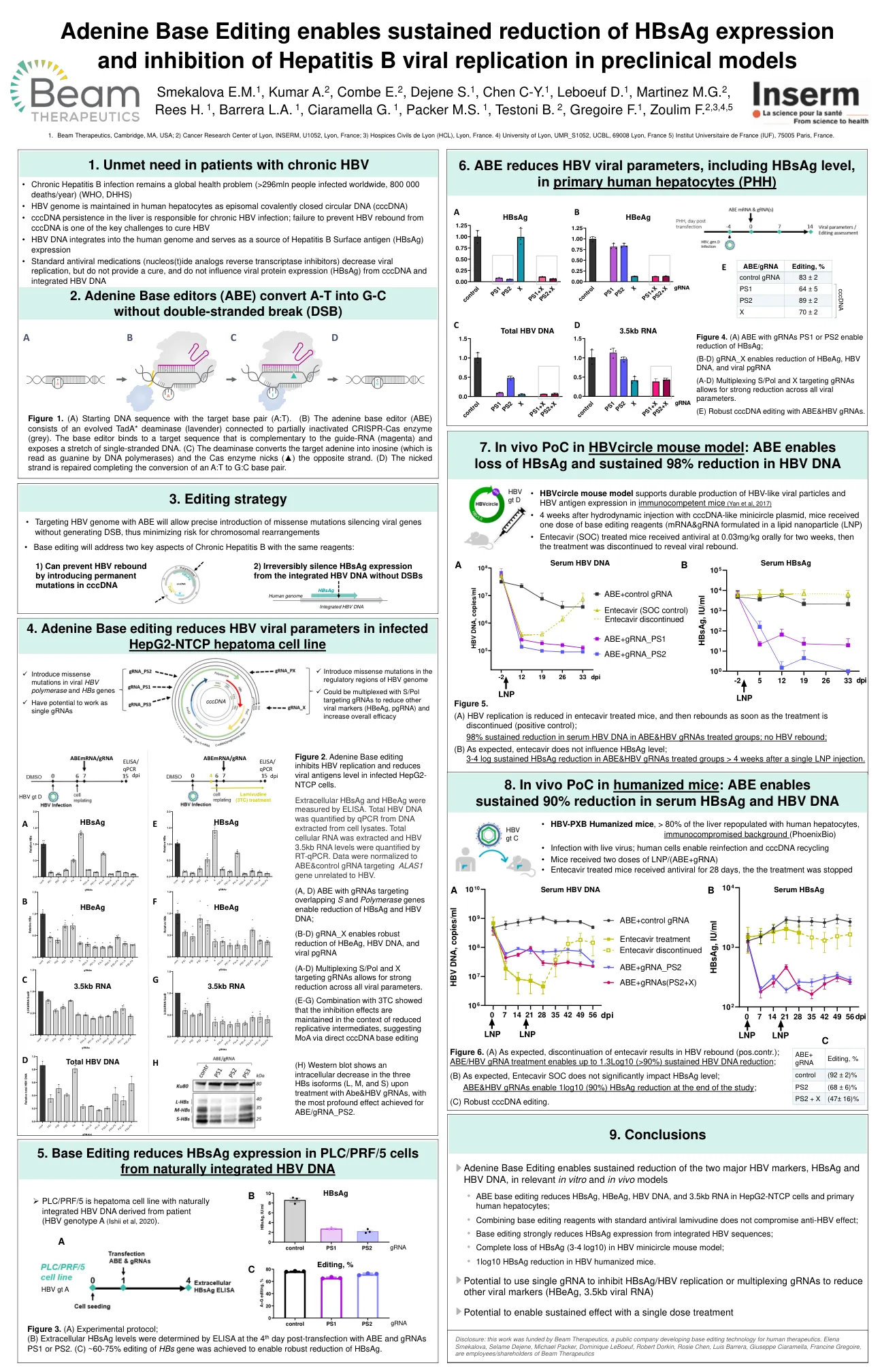
腺嘌呤碱基编辑能够在相关的体外和体内模型中持续降低两种主要的 HBV 标志物 HBsAg 和 HBV DNA
图 1. (A) 起始 DNA 序列,其中包含目标碱基对 (A:T)。(B) 腺嘌呤碱基编辑器 (ABE) 由进化的 TadA* 脱氨酶 (淡紫色) 和部分失活的 CRISPR-Cas 酶 (灰色) 组成。碱基编辑器与与向导 RNA (洋红色) 互补的目标序列结合,并暴露一段单链 DNA。(C) 脱氨酶将目标腺嘌呤转化为肌苷 (DNA 聚合酶将其读取为鸟嘌呤),Cas 酶切口 (▲) 另一条链。(D) 切口链被修复,完成从 A:T 到 G:C 碱基对的转换。