XiaoMi-AI文件搜索系统
World File Search SystemUTR
![arxiv:2502.13785v2 [q-bio.gn] 2025年3月11日](/simg/5\595a1c1f466cf12df861ef409dff5b847e21eabd.webp)
arxiv:2502.13785v2 [q-bio.gn] 2025年3月11日
基于mRNA的疫苗已成为药物工业的主要重点。mRNA的编码顺序以及未翻译区域(UTRS)可以强烈影响翻译效率,稳定性,降解和其他共同确定疫苗有效性的因素。但是,这些特性的光学mRNA序列仍然是一个复杂的挑战。逐步学习深度学习模型通常仅专注于编码区域优化,忽略UTR。我们提出了Helix-MRNA,这是一种基于结构化的状态空间和注意力混合模型,可应对这些挑战。除了第一次预训练外,第二个预训练阶段还使我们能够使用高质量的数据来专业。我们使用密码子分离对mRNA序列进行单核苷酸令牌化,从而确保了原始mRNA序列的先前生物学和结构信息不会丢失。我们的模型Helix-MRNA在分析UTR和编码区域属性方面优于现有方法。它可以比当前方法长6倍,同时仅使用现有基础模型的10%参数。其预测能力扩展到所有mRNA区域。我们开放码(https://github.com/ helicalai/helical)和模型权重(https://huggingface.co/ helical-ai helical-ai/helix-mrna)。

人类 T 细胞采用保守的 AU-
持久的 CD8 + T 细胞反应对于抵抗感染和肿瘤至关重要。促炎细胞因子 IFN- γ 是其中的关键效应分子。我们最近发现,在小鼠 T 细胞中,IFN- γ 的产生受到位于 Ifng mRNA 分子 3′ 非翻译区 (UTR) 的腺苷酸尿苷酸富集元素 (ARE) 的严格调控。ARE 的缺失导致活化 T 细胞中细胞因子产生时间延长,并增强抗肿瘤 T 细胞反应。在这里,我们研究了这些发现是否可以转化为原代人类 T 细胞。利用 CRISPR-Cas9 技术,我们从外周血来源的人类 T 细胞中的 IFNG 3′ UTR 中删除了 ARE 区域。ARE 的缺失使 T 细胞中的 IFNG mRNA 稳定,并支持更高比例的 IFN- γ 蛋白产生 T 细胞。重要的是,将 MART-1 T 细胞受体工程与 ARE-Del 基因编辑相结合表明,这也适用于抗原特异性激活 T 细胞。MART-1 特异性 ARE-Del T 细胞在响应表达 MART-1 的肿瘤细胞时,产生 IFN-γ 的 T 细胞百分比更高。综合起来,我们的研究表明,ARE 介导的转录后调控在小鼠和人类 T 细胞之间是保守的。此外,生成抗原特异性 ARE-Del T 细胞是可行的,这一特征可能用于治疗目的。

鉴定潜在的治疗靶标,以发展疾病管理中的新治疗策略
通过在1993年发现MicroRNA(miRNA),Victor Ambros1及其来自哈佛大学的群体为实现了研究领域的新里程碑做出了贡献。lin-4,其中包括与秀丽隐杆线虫的LIN-14 mRNA的3'未翻译区域(UTR)中的重复序列基序互补的序列。之后,Lin-4被视为蠕虫遗传学领域的发现。另一方面,直到发现第二个称为let-7的miRNA直到发现miRNA在包括人类在内的所有动物物种中都高度保守。

通过整合组学数据集进行 RNA 编辑的基因组鉴定及其在肝细胞癌中的临床意义
RNA 编辑是一种广泛使用的转录后机制,可在人类癌症中将单核苷酸变化引入 RNA。在本文中,我们对来自 Cancer Genome Atlas (TCGA) 的 373 个肝细胞癌 (HCC) 和 50 个相邻正常肝脏样本的整体 RNA 编辑谱进行了表征,并揭示了大多数编辑事件往往发生在少数具有中等编辑程度 (20-30%) 的样本中。此外,这些 RNA 编辑倾向于在蛋白质编码基因中发生 A 到 I 的 RNA 编辑,尤其是在 3′UTR 区域。考虑到 DNA 突变和 RNA 编辑之间的关联,我们的分析发现 RNA 编辑可能是 HCC 患者 HCC 风险基因 DNA 突变的互补事件。接下来,我们确定了 454 个与 HCC 相关的编辑位点,其中许多位于具有相同编辑模式的相同基因上。编辑的功能后果揭示了 2,086 个功能性编辑位点,并证明编码区域中的大多数编辑都是非同义变异。此外,我们的研究结果表明,3′UTR 区域的编辑往往会影响 miRNA 与靶标的结合,并且编辑程度似乎与基因表达呈负相关。最后,我们发现 46 个与 HCC 相关的编辑位点能够区分 HCC 患者的预后差异,表明它们具有临床相关性。总之,我们的研究结果强调 RNA 编辑是研究 HCC 机制和临床治疗的宝贵分子资源。

体内 CRISPR 筛选 mir- 的表型目标......
识别 miRNA 靶基因很困难,而确定哪些靶标在生物学上最重要则更加困难。我们设计了一种新策略,通过 CRISPR - Cas9 基因组编辑破坏秀丽隐杆线虫中每个预测的 miRNA 结合位点,来测试单个 microRNA - 靶标相互作用对表型的影响。我们开发了一种多重负选择筛选方法,其中对编辑的位点进行深度测序,并根据对破坏 miRNA 结合的突变的明显选择压力对候选位点进行优先排序。重要的是,我们的筛选是在突变动物体内进行的,这使我们能够研究生物体水平的表型。我们使用这种方法筛选了必需的 mir-35-42 家族的表型靶标。通过在所有预测靶标中生成 1130 个新的 3′UTR 等位基因,我们将 egl-1 确定为表型靶标,其去抑制部分表型复制了 mir-35-42 突变体表型,诱导了胚胎致死和低繁殖力。这些表型可以通过补偿性 CRISPR 突变来挽救,这些突变将 mir-35 重新定位到突变的 egl-1 3′UTR。这项研究表明,体内全生物体 CRISPR 筛选的应用具有加速发现非编码基因组中表型负调控元件的巨大潜力。
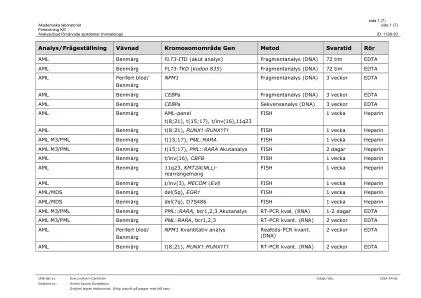
分析提供获得的疾病(血液学)
leukemiutringbenmärg/blod mylood面板*(ABL1,ANKRD26,ASXL1,ATRX,BCOR,BCOR,BCORL1,BRAF,CALR,CALR,CBL,CBL,CBL,CBL,CDKN2A,CDKN2A,CEBPA,CEBPA,CEBPA,CSF3R,CSF3R,CSF3R,CUX1,DDX41,DDX41,DNMT3A fbxw7, FLT3, GATA1, GATA2, GNAS, HRAS, Idh1, Idh2, Ikzf1, jak2, jak3, kdm6a, kit, kraas, kmt2a, mpl, myd88, NF1, Notch1 (INKLUSIVE 3´UTR), NPM1, NRAS, PDGFRA, PHF6, PPM1D, Pten, Ptpn11, Rad21, Runx1, Samd9, SAMDL9, Setbp1, SF3B1, SMC1A, SMC3, SRSF2, Stag2, Stat3, Stat5B, Tet2, TP53, U2AF1, WT1, ZRSR2, BTK, plcg2, terc) Div>

HSA-MIR-323A-3P充当肿瘤抑制因子,并靶向神经母细胞瘤细胞中的STAT3
背景:在过去几十年中进行的研究揭示了非编码microRNA(miRNA)在癌症发展和进展中的作用。染色体区域14q32中的几个miRNA(通常在癌症中通常删除的区域)与儿童癌症神经母细胞瘤的临床结果不良有关。与同一患者的预处理细胞相比,我们以前已经从该区域鉴定了该区域的miR-323A-3P在化学治疗治疗的神经母细胞瘤细胞中被下调。此外,在神经母细胞瘤肿瘤中,该miRNA在晚期4阶段疾病中被下调,与第1-2阶段相比在这项研究中,我们试图描述miR-323a-3p在神经母细胞瘤中的未知功能作用。方法:合成miRNA模拟物用于在神经母细胞瘤细胞系中过表达miR-323a-3p。研究了miR-323a-3p的功能作用,细胞活力测定,流量细胞术,反转录 - 定量聚合酶链反应,荧光素酶报道器分析和蛋白质印迹在神经母细胞瘤细胞系kelly,sh-sy5y和sh-sy5y和sk-sy5y和sk-n-be(2)-c(2)-c。结果:miR-323a-3p的异位表达导致凯利,sh-Sy5y和sk-n-be(2)-c的细胞活力显着降低,通过在所有细胞系中引起kelly,sh-Sy5y和sh-sy5y和凋亡中的G1细胞周期停滞。此外,在miR-323a-3p过表达时,降低了信号传感器的mRNA和蛋白质水平(STAT3)。结论:miR-323a-3p通过G1细胞周期停滞和凋亡抑制神经母细胞瘤细胞系的生长,而众所周知的癌基因STAT3是该miRNA的直接靶标。miR-323a-3p与Stat3的3'UTR的直接结合通过荧光素酶报道器测定在实验上验证,其中miR-323A-3P降低了来自全长Stat3 3'UTR荧光素酶报告剂的发光信号,但不是来自具有预测种子序列中突变的记者。

隔离,识别,分子表征
摘要本研究报告了奶牛场的流产,腹泻和牛奶生产急剧下降。该农场通常用进口疫苗接种了针对BVDV的疫苗,其中含有典型的Pestiviruses菌株(BVDV-1和BVDV-2)。从流产的母牛和显示持续性腹泻的奶牛中收集了总共13个血清样品,5个阴道排放样品和5个粪便样品。使用PCR筛选所有样品的潜在微生物原因(病毒或细菌)。在测试的23个样品中,只有一个阴道放电样品在预期的288 bp下产生了阳性的PCR结果。设计的引物是对基于5'-UTR的RTPCR测定法的高灵敏度,用于检测Pestiviruses。将PCR产品发送进行序列分析,并将结果提交给GenBank登录号#OR425033,并设计为GERD/VSVRI/PESTI-GIRAFFE/2022。然后通过三个连续的盲传中成功地在MDBK细胞中成功分离并传播该病毒。在病毒后接种后2-3天观察到了一种明显的细胞质效应(CPE),其特征是感染后72小时,其特征是液泡,细胞舍入和簇形成。pcr均在每个段落上进行,并以预期的大小给出了一个特定的频带。通过序列比对和系统发育分析的进一步分析表明,分离株与Pestivirus长颈鹿密切相关,尤其是Pestivirus PG-2。这标志着该菌株在埃及的检测,隔离和表征的第一个记录。因此,这种流行是由埃及记录的新引入的菌株引起的。因此,进口的疫苗无法提供保护,需要更新当地的疫苗以包括此Pestivirus菌株。关键字:Pestivirus PG-2,PNS,MDBK,5`UTR,CPE,系统发育分析,PCR,BDV,

多动物模型研究揭示了神经可塑性和伤害性基因的突变与饮酒过量有关
结果:我们的第一个至关重要的发现是,除了引起翻译变化的变体外,与饮酒前的饮酒相关的主要遗传变化也称为“沉默突变”和3'未翻译区域(3'UTR)中的突变。这些都没有改变所翻译的氨基酸序列,而是影响基因转录的速率和构象,包括改变基因疗效的稳定性和翻译后事件。这一发现提倡在人类基因组研究中重新聚焦基因效能感的变化。在确定的关键本体论中是“疼痛的伤害感受或感觉感知”,它不仅包含伤害感受(ARRB1,CCL3,EPHB1),而且还伴有钠(SCN1A,SCN1A,SCN2A,SCN2B,SCN2B,SCN3A,SCN3A,SCN7A,SCN7A),SCN7A),SCN99A(SCN9A9A)(kc N9aa)(KC)和POTASS(kc)。

1 LeishGEdit 工具箱的 Bar-seq 策略 2
LeishGEdit 引物设计流程为基因组中每个给定的 ORF 设计了总共六个引物序列,以实现 CRISPR-Cas9 基因编辑,从而允许用大量可用标签标记感兴趣基因的 N 端或 C 端,并删除 ORF 的两个等位基因(图 1B 和 C)。设计了两个 sgRNA 引物,一个靶向目标基因的 5'UTR,一个靶向 3'UTR。sgRNA 引物由 T7 RNAP 启动子序列、20 nt sgRNA 靶序列(用于在感兴趣的位点引入 DSB)和 20 nt 与 CRISPR-Cas9 主链序列的重叠组成,从而允许通过 PCR 生成 sgRNA 模板。需要一个包含整个 sgRNA 主链序列 [20] 的额外通用引物来扩增两个 sgRNA。四种引物专为 pPLOT 和 pT 质粒扩增而设计,可以以不同的组合使用 82 来产生供体 DNA。这些引物包括:上游正向引物 (#1)、83 上游反向引物 (#2)、下游正向引物 (#3) 和下游反向引物 (#4)。可以选择性地设计 84 额外引物,以允许使用从 pPOT 质粒模板扩增的供体 85 构建体进行 CRISPR-Cas9 介导的基因编辑 [21]。供体 DNA 引物包含紧邻 sgRNA 靶序列及其 PAM 位点的 30 nt HF 序列 86,以及与 pT、pPLOT 和 pPOT 质粒兼容的引物结合位点 87。虽然上游正向引物和下游 88 反向引物位置始终根据所选的 sgRNA 而变化,但上游反向引物 89 (#2) 和下游正向引物 (#4) 针对每个基因设计在相同的位置。90