XiaoMi-AI文件搜索系统
World File Search Systemactin

大学'di siena dipartimento di Medicina Molecolare e Dello Sviluppo dottorato dotorato di ricerca in Medicina molecolare ciclo ciclo ciclo xxxv coordinatore c
图3形成肌肉纤维的收缩元件的示意图。由肌动蛋白和辅助蛋白肌动蛋白和肌钙蛋白组成的细丝锚定在Z盘上,Z盘主要由棒状肌动蛋白结合蛋白α-肌动蛋白形成。肌球蛋白形成的厚细丝由结构蛋白肌瘤锚定在M系的水平。其他基本蛋白质是钛,它像分子弹簧一样连接到Z-二烟的厚细丝,在肌肉发育过程中涉及肌动蛋白组装的Nebulin和smbisturin,它通过与Sank1.5的相互作用将收缩仪与SR膜连接到SR膜。图像改编自(Gokhin&Fowler,2011年)。

大脑通讯 AIN 通讯
树突棘中肌动蛋白细胞骨架动力学的调节对于学习和记忆形成至关重要。因此,肌动蛋白细胞骨架通路缺陷是多种脑部疾病(包括阿尔茨海默病)的生物学特征。本文,我们描述了一种由环化酶相关蛋白 2 控制的新型突触机制,该蛋白是结构可塑性现象所必需的,在阿尔茨海默病中完全被破坏。我们报告称,通过其 Cys 32 形成环化酶相关蛋白 2 二聚体对于环化酶相关蛋白 2 与辅酶蛋白结合以及肌动蛋白周转非常重要。Cys 32 依赖性环化酶相关蛋白 2 同源二聚化和与辅酶蛋白的结合由长期增强作用触发,并且是长期增强诱导的辅酶蛋白易位到棘突、棘突重塑和突触传递增强所必需的。这种机制在阿尔茨海默病患者和 APP/PS1 小鼠的海马中特别受到影响,但在额上回中没有受到影响,其中环化酶相关蛋白 2 下调,环化酶相关蛋白 2 二聚体突触水平降低。值得注意的是,阿尔茨海默病患者的脑脊液中环化酶相关蛋白 2 水平显著升高,但在额颞叶痴呆患者中没有升高。在阿尔茨海默病海马中,cofilin 与环化酶相关蛋白 2 二聚体/单体的关联发生改变,并且 cofilin 在脊柱中异常定位。总之,这些结果为阿尔茨海默病中存在缺陷的结构可塑性机制提供了新的见解。

三年完全资助的博士职位
三年全额资助的博士职位(研究生院 - 化学,生物学和健康)地点:格勒诺布尔神经科学研究所,“神经循环骨骼动力学和结构”团队,CheminFortunéFerrini,38700 La Tronche,法国,法国。标题:探索Tau在细胞粘附动力学中的作用:Tau,细胞骨架和局灶性粘附摘要之间的相互作用:与神经变性相关的蛋白质Tau最近成为癌症的预后因素。然而,连接tau和细胞转化的分子机制仍然很少了解。作为tau是微管和肌动蛋白细胞骨架的关键参与者,其错误调节可能会改变几个事件,包括细胞粘附,迁移和转移。的确,最近的研究强调了局灶性粘附途径与tau功能障碍之间的联系。该项目旨在探讨Tau的表达和/或修饰如何通过微管和/或肌动蛋白网络的破坏,涉及肿瘤进展的过程来影响局灶性粘连。By combining in cellulo and in vitro studies, the PhD candidate will pursue two main objectives: 1/ to correlate, in cell models, tau- mediated cytoskeleton organisation with focal adhesion assembly and distribution 2/ to reconstitute cytoskeleton properties with purified proteins and investigate how tau proteins affect the microtubule/actin interplay known to occur at the leading edge of migrating cells.总体而言,结果应强调tau蛋白在肿瘤发生错误调节的机制中的作用,并有助于了解神经退行性疾病与癌症之间的联系。

重组大鼠微管相关蛋白tau(mapt ...
tau蛋白是一种由MAPT基因编码的高度可溶的微管相关蛋白(MAP)。tau蛋白是一种基本蛋白。作为地图家族的成员,tau蛋白主要作用于轴突的远端,以维持微管的稳定性和柔韧性。tau蛋白与微管蛋白相互作用以稳定微管,同时驱动微管内的小管蛋白组装。tau蛋白通过异构化和磷酸化控制微管的稳定性。tau蛋白参与调节轴突运输和核功能以保护DNA完整性。与肌动蛋白细胞骨架相互作用以促进肌动蛋白丝的形成;并通过与FYN相互作用来调节NMDA受体信号通路。tau的磷酸化受许多激酶的调节,包括PKN,丝氨酸/苏氨酸激酶,其活化会导致微管组织破坏。高磷酸化TAU在神经元中的积累会引起神经原纤维变性,这与各种神经退行性疾病(如AD和PD)有关。
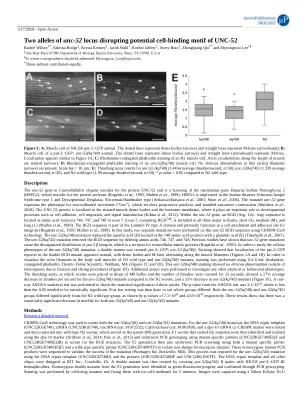
unc-52 基因座的两个等位基因破坏了潜在的细胞......
图 1:A:NK358 pat-3::GFP 动物的肌肉细胞。虚线代表致密体(箭头),直线代表 M 线(箭头);B:pat-3::GFP; unc-52(kq748) 动物的肌肉细胞。虚线代表致密体(箭头),直线(箭头)代表 M 线。定位看起来与图 1A 相似;C:N2 肌肉细胞的罗丹明偶联鬼笔环肽染色。沿肌肉长度的肌动蛋白细胞骨架被染色(箭头);D:unc-52(kq748) 肌肉细胞的罗丹明偶联鬼笔环肽染色。细(肌动蛋白)丝(箭头)中没有明显异常。比例尺 = 10 µm。; E:unc-52 (kq748)(平均每秒 1.4454 次冲击,n=50)、unc-52(kq745)(平均每秒 1.339 次冲击,n=50)和 N2 野生型(平均每秒 1.99 次冲击,n=50)的冲击试验结果。 * 与 N2 野生型相比,p 值 < 0.05。

LPAR1重组单克隆抗体
相关性溶血磷脂酸 (LPA) 受体 (PubMed:9070858, PubMed:19306925, PubMed:25025571, PubMed:26091040)。在肌动蛋白细胞骨架重组、细胞迁移、分化和增殖中发挥作用,从而有助于对组织损伤和感染因子的反应。通过异源 G 蛋白的 G(i)/G(o)、G(12)/G(13) 和 G(q) 家族激活下游信号级联。信号抑制腺苷酸环化酶活性并降低细胞 cAMP 水平 (PubMed:26091040)。信号传导触发细胞质 Ca(2+) 水平的增加 (PubMed:19656035, PubMed:19733258, PubMed:26091040)。激活 RALA;这导致磷脂酶 C (PLC) 的激活和肌醇 1,4,5-三磷酸的形成 (PubMed:19306925)。信号传导介导下游 MAP 激酶的激活 (通过相似性)。有助于调节细胞形状。促进神经元细胞中肌动蛋白细胞骨架的 Rho 依赖性重组和神经突回缩 (PubMed:26091040)。促进 Rho 的激活和肌动蛋白应力纤维的形成 (PubMed:26091040)。通过激活 RAC1 促进迁移细胞前缘板状伪足的形成(通过相似性)。通过其作为溶血磷脂酸受体的功能,在趋化性和细胞迁移中发挥作用,包括对损伤和创伤的反应(PubMed:18066075,PubMed:19656035,PubMed:19733258)。通过与 CD14 相互作用,在引发对细菌脂多糖 (LPS) 的炎症反应中发挥作用。促进对溶血磷脂酸的细胞增殖。正常骨骼发育所必需的。可能在成骨细胞分化中发挥作用。正常大脑发育所必需的。成人齿状回中新形成的神经元正常增殖、存活和成熟所必需的。在疼痛感知和神经性疼痛的引发中发挥作用(通过相似性)。

RHOA/RHOB的高表达比与基底样乳腺肿瘤的迁移和侵入性特性有关
基底样乳腺癌是最具侵略性的癌症之一,仍然没有有效的靶向治疗方法。为了鉴定新的治疗靶标,我们在八个乳腺癌细胞系上进行了mRNA-SEQ。在基础样肿瘤中过表达的基因中,我们专注于RhoA和RhoB基因,该基因编码已知在肌动蛋白细胞骨架中起作用的小GTPases,从而允许细胞迁移。QRT-PCR和Western印迹用于表达研究。通过伤口愈合和Boyden Chambers分析分析了迁移和侵入性特性。通过荧光肌动蛋白标记评估应力纤维的形成。Rho siRNA,小型抑制剂Rhosin处理和BRCA1转染以研究RHO和BRCA1蛋白的作用。我们表明,RhoA的强烈表达和RHOB的低表达与乳腺癌的基础样亚型有关。降低RhoA表达可降低基底样细胞系的迁移和侵袭能力,同时降低RHOB表达增加了这些能力。Rhosin是RhoA的抑制剂,也可以减少基底样细胞系的迁移。RHO蛋白参与了应激纤维的形成,这是迁移细胞中发现的肌动蛋白细胞骨架的构象:RhoA表达的抑制降低了这些纤维的形成。这些结果表明,Rho蛋白是基底样和BRCA1突变乳腺癌的潜在治疗靶标,因为迁移和获得间充质特性是这些具有高转移性潜力的肿瘤的关键功能途径。brca1是一种基础样肿瘤中经常失活的基因,似乎在这些肿瘤中RhoA和RhoB的差异表达中起作用,因为在BRCA1突变的基底样细胞系中BRCA1表达的恢复RhoA的表达降低了RhoA的表达和RHOB的表达,并增加了迁移能力的表达。
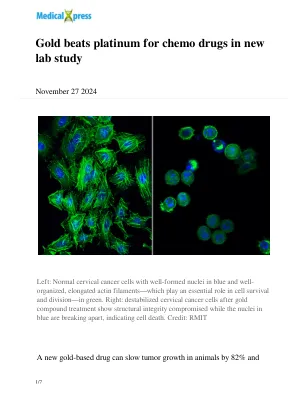
新实验室研究表明黄金在化疗药物中的应用优于铂金
左图:正常宫颈癌细胞,细胞核结构良好(蓝色),肌动蛋白细丝(绿色)排列整齐,肌动蛋白细丝在细胞存活和分裂中起着至关重要的作用。右图:不稳定的宫颈癌细胞经金化合物处理后,结构完整性受损,细胞核(蓝色)分裂,表明细胞死亡。图片来源:RMIT

Wiskott Aldrich 综合征蛋白调节人类髓系细胞的非选择性自噬和线粒体稳态
摘要 肌动蛋白细胞骨架调节剂 Wiskott Aldrich 综合征蛋白 (WASp) 与先天性鼠免疫细胞中自噬-炎症小体轴的维持有关。本文表明,WASp 缺乏与雷帕霉素诱导的自噬体形成和运输到原代人类单核细胞衍生巨噬细胞 (MDM) 中的溶酶体受损有关。临床基因治疗后,体外和 WAS 患者体内的 WASp 重建可恢复自噬通量,并且依赖于肌动蛋白相关蛋白复合物 ARP2/3。作为选择性自噬的模型,用 CCCP 诱导线粒体损伤也揭示了 WASp 在形成隔离肌动蛋白笼和维持线粒体网络完整性方面具有新的 ARP2/3 依赖性作用。此外,WAS 患者 MDM 中的线粒体呼吸受到抑制,并且在应激时无法达到正常的最大活动,表明存在严重的内在代谢功能障碍。总之,我们提供了人类 WASp 在自噬过程和免疫代谢调节中发挥的新重要作用的证据,这可能在机制上促成了复杂的 WAS 免疫表型。
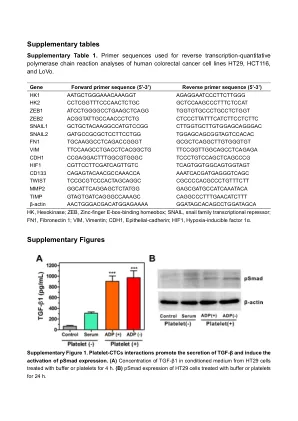
补充表
ctctttctctctc nail1 gctgctgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtggtggggggghts snal2 TGCAAGCCTCGGGGTGTGTGGTGGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGCCCCCCCCCCCCCGCTGGTGGCTGGTGTGTGTGTGGTGH1 CDH1 CDGGTGGGGC TCCTGTCCAGCCCCGCCGCCCGCGCGCGCCGCGGS CGTTCTCTCTGTGTC TCAGTGGGGGGGGGTGGTGTGTGTGTs CD133 CAGTACCAACCAAACCIGITCIGITITIs GAGCIGITING TIMP GTAGGGGGGGGGGGGGGGGGGCAGC caggccttttgttttttttttttttttttttttttsβ-肌动蛋白actghtgacity gtatagcagcatgityctctttctctctc nail1 gctgctgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtggtggggggghts snal2 TGCAAGCCTCGGGGTGTGTGGTGGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGCCCCCCCCCCCCCGCTGGTGGCTGGTGTGTGTGTGGTGH1 CDH1 CDGGTGGGGC TCCTGTCCAGCCCCGCCGCCCGCGCGCGCCGCGGS CGTTCTCTCTGTGTC TCAGTGGGGGGGGGTGGTGTGTGTGTs CD133 CAGTACCAACCAAACCIGITCIGITITIs GAGCIGITING TIMP GTAGGGGGGGGGGGGGGGGGGCAGC caggccttttgttttttttttttttttttttttttsβ-肌动蛋白actghtgacity gtatagcagcatgity