XiaoMi-AI文件搜索系统
World File Search Systemcas9

Cas9 融合可实现精确的体内编辑
目前的 Cas9 试剂可以高度特异性地靶向基因组位点。然而,当用于敲入时,靶向结果本质上是不精确的,通常会导致非预期的敲除而不是预期的编辑。这将基因组编辑的应用限制在离体方法中,其中可能进行克隆选择。在这里,我们描述了一种使用迭代高通量体外和高产量体内测定的工作流程,以评估和比较 CRISPR 敲入试剂在编辑效率和精度方面的性能。我们测试了 Cas9 和 DNA 供体模板变体的组合,并确定 Cas9-CtIP 与原位线性化供体在细胞系和小鼠脑体内显示出成倍的编辑精度增加。通过迭代此过程,我们生成了新的化合物融合,包括 eRad18-Cas9-CtIP,其性能进一步成倍增加。继续利用该平台开发精确编辑试剂有望在模型生物中直接进行体内敲入,并有望用于未来的靶向基因疗法。
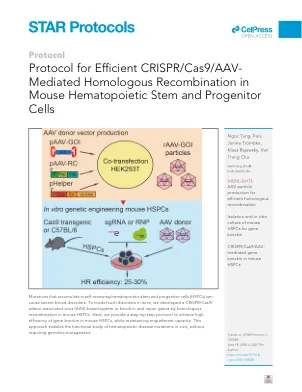
高效 CRISPR/Cas9/AAV 协议
1. 为了在小鼠 HSPC 中实现有效的同源重组 (HR) 事件,需要具有高编辑效率的特定单向导 RNA (sgRNA)。我们使用 CrispRGold 程序 ( https://crisprgold.mdc-berlin.de ) 来设计特定的 sgRNA 并预测潜在的脱靶 ( Chu et al., 2016a )。每个目标序列应设计几个特定的 sgRNA。必须通过使用 T7 内切酶 I 测定 ( Guschin et al., 2010 ) 测量错配的 DNA 异源双链体以及对至少 2 种主要血细胞类型(例如 B 细胞和 T 细胞)的 PCR 产物进行 Sanger 测序来验证所有 sgRNA 的编辑效率。可以从 IDT、Synthego 或其他供应商处订购化学修饰或未修饰形式的 sgRNA。 2. 供体模板的最佳设计对于小鼠 HSPC 中的高效 HR 至关重要。供体模板包括 5'、3' 同源臂和所需的修饰基因序列。同源臂的长度取决于目标序列的特异性,每个同源臂由 600 到 2000 bp 组成。AAV 基因组的包装能力是设计供体模板的一个限制,因为基于 AAV 的供体模板的最大长度不应超过 4.5kb。如果没有使用报告基因,则应通过引入可用于量化 HR 效率的沉默突变将限制性酶识别位点添加到修饰的基因序列中。3. 为了通过 PCR 扩增和测序量化目标位点中的 HR 和非同源末端连接 (NHEJ) 事件,必须在外部设计正向或反向引物,或两者

在桉树中实施 CRISPR/Cas9 技术...
1 植物科学研究实验室,图卢兹第三大学,CNRS,UPS,UMR 5546,24 Chemin de Borde Rouge,31320 Castanet-Tolosan,法国; ying.dai@lrsv.ups-tlse.fr (YD); annabelle.dupas@lrsv.ups-tlse.fr (广告); luciano.medina@lrsv.ups-tlse.fr (LM); nils.blandel@lrsv.ups-tlse.fr (注意); san-clemente@lrsv.ups-tlse.fr(HSC); ladouce@lrsv.ups-tlse.fr(荷兰); mounet@lrsv.ups-tlse.fr(调频); grima@lrsv.ups-tlse.fr (JG-P.) 2 UMR 990,水果基因组学和生物技术,图卢兹大学,INP-ENSA 图卢兹,Avenue de l'Agrobiopole,31326 Castanet-Tolosan,法国; guojian.hu@etu.ensat.fr 3 TBI,图卢兹大学,CNRS,INRAE,INSA,31400 图卢兹,法国; Myriam.Badawi@univ-lemans.fr(MB); hernandg@insa-toulouse.fr (GH-R.) 4 海洋分子健康实验室,MMS EA2160 勒芒大学,72085 勒芒,法国 * 通讯地址:wang@lrsv.ups-tlse.fr

Cas9 介导的 β-catenin 突变校正和...
目的:结直肠癌 (CRC) 是导致癌症死亡和发病率的主要原因之一。迫切需要找到对抗 CRC 的策略。APC 或 β -catenin 的驱动基因突变在 CRC 的发生和进展中起重要作用。在本研究中,我们联合应用 CRISPR/Cas9-sgRNA 系统和单链寡脱氧核苷酸 (ssODN) 作为模板来纠正结肠癌细胞系 HCT-116 中存在的 β -catenin 的杂合 Δ TCT 缺失突变。该方法为癌症的基因治疗提供了一种潜在的策略。方法:构建 Cas9/β -catenin-sgRNA-eGFP 共表达载体并与 ssODN 共转染到 HCT-116 细胞中。通过 FACS 分选突变校正的单细胞克隆,并通过 TA 克隆和 DNA 测序进行判断。通过实时定量PCR、Western印迹、CCK8、EDU染色和细胞接种克隆检测CRISPR/Cas9介导的校正效果。此外,还分析了裸鼠异种移植瘤中细胞克隆衍生肿瘤的生长情况。结果:CRISPR/Cas9介导的β-catenin突变校正导致TCT序列的存在和Ser45处磷酸化β-catenin的重新表达,从而恢复了磷酸化β-catenin的正常功能,包括减少核β-catenin的运输和下游c-myc、survivin的表达。在β-catenin突变校正的细胞中观察到细胞生长显著减少。移植了突变校正的HCT-116细胞的小鼠的肿瘤大小明显小于未校正的异种移植瘤。结论:本研究数据表明,通过 CRISPR/Cas9 和 ssODN 的组合来纠正驱动突变可以极大地改善癌细胞系的生物学行为,表明该策略在癌症基因治疗中具有潜在的应用价值。关键词:CRISPR/Cas9、ssODN、靶向基因编辑、β-catenin、结肠癌

转化生物医学中的 CRISPR/Cas9 技术
摘要 成簇的规则间隔短回文重复序列 (CRISPR) - RNA 引导的 Cas9 内切酶系统为包括人类在内的多种哺乳动物物种的精确基因组编辑提供了一种快速有效的方法。CRISPR/Cas9 技术允许通过进行删除、插入或 DNA 供体指导的精确序列修饰,在一个主要步骤中对所选基因的位点特定位置进行修饰。Cas9 与序列特异性引导 RNA 形成核蛋白复合物,以在互补 DNA 靶中产生双链断裂。此外,双链断裂修复机制可导致预期的基因修饰。CRISPR/Cas9 系统是一种广泛用于基因组修饰、编辑和其他生物技术应用的技术,例如功能注释、用于可视化特定基因组位点的系统和基因的转录控制。CRISPR/Cas9 介导的实验动物基因组操作有助于理解基因功能,并已成为一种模拟人类疾病的流行方法。此外,CRISPR-Cas9 系统在人类基因中的应用日益广泛,成为一种用于人类疾病分子鉴定和治疗的极其强大的技术。在这篇综述中,我们介绍了 CRISPR/Cas9 技术的基本原理及其在转化生物医学中的应用的最新进展。

Guide-it CRISPR/Cas9 基因编辑
◀ Cas9多克隆抗体检测灵敏度(Western blot)(泳道3: 0.15 ng, 4: 0.30 ng, 5: 0.625 ng, 6: 1.25 ng, 7: 2.5 ng, 8: 5.0 ng, 9: 10.0 ng, 10: 20.0 ng)

CRISPR/Cas9 靶向多药耐药性
图 2. Western Blot 膜图像(WT:野生型,KO-mecA:mecA 基因抑制菌株)如图 3 所示,与野生型 (WT) 相比,KO 菌株中 PBP2a 表达显著减少 70%,进一步验证了成功破坏了甲氧西林耐药性。这些发现不仅展示了 CRISPR 技术在实现有针对性的基因改造方面的效力,而且还强调了其在基因和表型水平上解决抗生素耐药性的转化潜力。调节关键耐药基因表达的能力有望推动针对多药耐药病原体的精准治疗。

theranostics a genome act crispr/cas9屏幕...
肽受体放射性核素疗法(PRRT)使用177个神经内分泌肿瘤(NET)的177 lutetium-dota-crottreotate(Lutate)现在在许多国家可以使用的批准治疗方法,尽管原发性或次要抵抗力继续限制其有效性或耐用性。我们假设,全基因组CRISPR/CAS9筛查将确定对黄体和基因靶标的反应的关键介体,这可能为净患者提供新型组合疗法的机会。方法:我们在露酸盐处理的细胞中使用了全基因组CRISPR-CAS9筛选,以鉴定影响细胞对鲁丁的敏感性或抗性的基因。命中通过单基因敲除验证。耐酸性细胞,以确认露丝的摄取和保留率,并持续生长抑素受体2(SSTR2)表达。基因敲除赋予黄酸盐敏感性的基因敲除,通过使用特定抑制剂和体内分析这些抑制剂与黄体结合使用的疗效,进一步表征了药理敏感性。结果:CRISPR-CAS9屏幕确定了对PRRT的耐药性和敏感性的几个潜在目标。两个基因敲除在体外赋予了放光抗性的基因敲除,ARRB2和MVP具有与Lutate结合和保留相关的潜在机制,分别对DNA破坏修复(DDR)途径的调节。屏幕表明,可以通过在DDR途径中涉及多种基因的损失来赋予对鲁酸酯治疗的敏感性,而非同源末端结合(NHEJ)的基因丧失是最致命的。通过基因丧失或通过两个不同抑制剂抑制键NHEJ基因PRKDC(DNA-PK)的丧失导致细胞在暴露于细胞时的生存率显着降低。在SSTR2阳性携带的小鼠中,Nedisertib(DNA-PK特异性抑制剂)和黄体的组合产生了对肿瘤生长的更强控制和与单独使用的肿瘤相比的生存率。结论:DDR途径对于传感和修复辐射诱导的DNA损伤至关重要,我们的研究表明,DDR途径的调节可能涉及对PRRT的耐药性和敏感性。此外,使用DNA-PK抑制剂与Lutate PRRT结合使用显着提高了治疗在临床前模型中的疗效,从而提供了进一步的证据证明该组合的临床功效。

crispr/cas9指示OSGA20OX2的诱变
摘要:在大米中,半弱SM是最需要的特征之一,因为它促进了更好的产量和耐药性。Here, semi-dwarf rice lines lacking any residual transgene-DNA and o ff -target e ff ects were generated through CRISPR / Cas9-guided mutagenesis of the OsGA20ox2 gene in a high yielding Basmati rice line, and the isobaric tags for relative and absolute quantification (iTRAQ) strategy was utilized to elucidate the proteomic changes in mutants.结果表明吉布林林(GA 1和GA 4)水平降低,植物高度(28.72%)和叶叶长度,而所有其他特征保持不变。OSGA20OX2表达得到了高度抑制,突变体表现出降低的细胞长度,宽度,并通过外源性GA 3处理恢复其植物高度。野生型和纯合突变系(GXU43_9)的比较蛋白质组学分别显示了588种蛋白质的水平,分别是273个上调和315个下调的水平。鉴定出的差异表达的蛋白质(DEP)主要富含碳代谢和固定,糖酵解 /糖糖异生,光合作用和氧化磷酸化途径。与生长调节因素(GRF2,GRF7,GRF9,GRF9,GRF11和GRF11)和GA(Q8RZ73,Q8RZ73,Q9AS97,Q69197,Q69VG1,Q69VG1,Q8LNJ6,Q8LNJ6,q8lnj6,q8lnj6,qy8lnj6,qy8lnj6,q55,在突变系中,脱离应激抗应激的蛋白5(ASR5)和脱落酸受体(PYL5)上调。我们将CRISPR / CAS9与蛋白质组学筛选整合为快速评估CRISPR实验结果的最可靠策略。

重组NLS-SPCAS9-NLS(CAS9)核酸酶解决方案
产品描述:Akron的重组NLS-SPCAS9-NLS(CAS9)核酸酶解决方案均遵循所有相关的辅助材料指南。下游纯化过程使用多步矫正方法,不使用亲和力标签,以最大程度地减少外源杂质,并确保提供高度纯化和活性的物质以进行进一步的制造应用。Akron的Cas9核酸酶是单链,162 kDa,非糖基化的DNA核酸内切酶,在大肠杆菌中表达。氨基酸序列来自链球菌链球菌,蛋白质结构已通过在蛋白质的N-和C-末端添加核定位序列(NLS)改变了蛋白质结构。将NLS添加到蛋白质结构中确保您的RNP复合物有效地导入细胞核。