XiaoMi-AI文件搜索系统
World File Search Systemcd8
naveni®CD8/MHC-I ATTO647N
图像:人类扁桃体组织中的CD8/MHC-I相互作用,与CD8/MHC-I ATTO647N检测到的红色相互作用,与CD3(CYAN)和核染色(DAPI)

INBRX- 120,一种靶向 CD8α 的失谐 IL
摘要 背景 作为淋巴细胞增殖和活化的主要驱动因素,白细胞介素 2 (IL-2) 是抗肿瘤反应的关键介质。尽管在部分患者中表现出良好的活性,但 IL-2(阿地白介素)的更广泛治疗效用受到严重的剂量限制性毒性、免疫抑制调节性 T 细胞的扩增和较差的药代动力学 (PK) 特征的阻碍。最近的工程改造努力,包括非 α IL-2 变体,已经降低了毒性特征,但尚未在更广泛的患者群体中诱导有意义的抗肿瘤活性。方法我们设计了 INBRX-120,一种 CD8 α 靶向 Cisleukin™ 分子,由亲和力调节的 IL-2 (IL2-x) 通过效应沉默的 Fc 结构域连接到两个高亲和力 CD8 α 特异性单域抗体组成。为了证明这种巨大的亲和力差异能够使 IL- 2 顺式信号传导专门作用于表达 CD8 α 的杀瘤效应细胞群,我们在体外测试了 INBRX-120 对靶细胞扩增、活化和抗肿瘤活性的影响。在同源小鼠模型中单独或与程序性细胞死亡蛋白 1 (PD-1) 阻断剂联合使用评估了体内抗肿瘤功效。在非人类灵长类动物中进行了临床前安全性以及药效学 (PD) 和 PK 分析。结果 INBRX-120 有效扩增和增强了 CD8 T 细胞和自然杀伤细胞对肿瘤细胞的细胞毒能力,而不会在体外和体内影响调节性 T 细胞。在同源小鼠模型中,INBRX-120 替代品单独使用和与 PD-1 阻断剂联合使用均表现出安全、有效和持久的抗肿瘤功效。在非人类灵长类动物中,INBRX-120 扩增并激活了表达 CD8 α 的效应细胞,显示出良好的 PK 特性,并且在剂量高达 1 mg/kg 时耐受性良好。结论通过其对表达 CD8 α 的效应细胞的独特顺式信号传导活性,INBRX-120 克服了基于 IL-2 的疗法的主要局限性,并有效地利用了 IL-2 强大的内在抗肿瘤活性。这种新颖的治疗策略有望实现更安全的临床活性,可在更广泛的各种癌症适应症患者中诱导有意义的抗肿瘤疗效。

使用 DART-AAV 进行体内基因传递,靶向 CD8
体内基因治疗面临的最大挑战之一是载体介导高度选择性的基因转移到特定治疗相关细胞群中。我们在此介绍 DARPin 靶向 AAV(DART-AAV),展示针对人类和鼠 CD8 的 DARPin。将 DARPin 插入 AAV2 和 AAV6 衣壳蛋白 1(VP1)的 GH2/GH3 环中,可实现对 CD8 阳性 T 细胞的高选择性,同时基因传递活性不受影响。值得注意的是,衣壳核心结构未发生改变,突出的 DARPin 可检测到。在复杂的原代细胞混合物中,包括供体血液或小鼠全身注射,CD8 靶向 AAV 在选择性、靶细胞活力和基因转移率方面远远优于未改造的 AAV2 和 AAV6。在体内,将单个载体注射到经过条件化的人源化或免疫功能正常的小鼠中,可击中高达 80% 的活化 CD8+ T 细胞。虽然在非活化条件下基因转移率显著降低,但在将 Cre 递送到指示小鼠中时,仍然可以检测到 CD8+ T 细胞中的选择性基因组修饰。在两种小鼠模型中,CD8+ T 细胞的选择性接近绝对,但肝脏的靶向性极强。本文描述的 CD8-AAV 扩展了免疫学研究和体内基因治疗选择的策略。

LEGENDplex™ 人类 CD8/NK 检测组标准 V02
741188 (LEGENDplex™ 人 CD8/NK 组检测抗体 V02) 741213 (LEGENDplex™ 缓冲液套装 R) 741190 (LEGENDplex™ 人 IL-17A 捕获珠 A3, 13X) 740268 (LEGENDplex™ 人 IL-2 捕获珠 A4, 13X) 740269 (LEGENDplex™ 人 IL-4 捕获珠 A5, 13X) 740270 (LEGENDplex™ 人 IL-10 捕获珠 A6, 13X) 741191 (LEGENDplex™ 人 IL-6 捕获珠 A7, 13X) 741192 (LEGENDplex™ 人 TNF-α 捕获珠 A10, 13X) 741193 (LEGENDplex™ 人 sFas捕获珠 B2,13X) 741194(LEGENDplex™ 人 sFasL 捕获珠 B3,13X) 741195(LEGENDplex™ 人 IFN-γ 捕获珠 B4,13X) 740277(LEGENDplex™ 人颗粒酶 A 捕获珠 B5,13X) 741196(LEGENDplex™ 人颗粒酶 B 捕获珠 B6,13X) 740279(LEGENDplex™ 人穿孔素捕获珠 B7,13X) 740280(LEGENDplex™ 人颗粒溶素捕获珠 B9,13X)
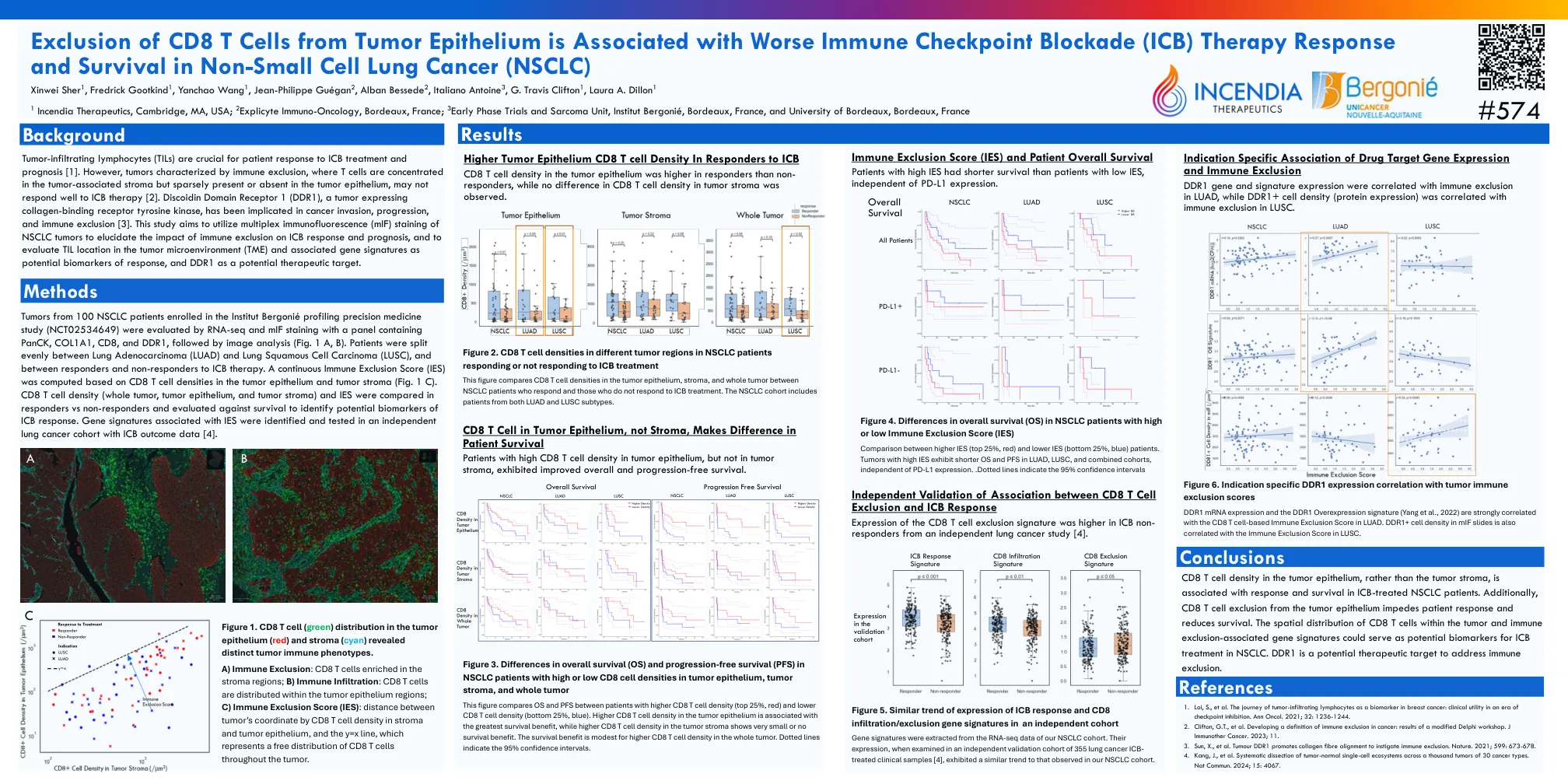
排除CD8 T细胞从肿瘤上皮中排除在...
通过RNA-SEQ和MIF染色评估了来自100名NSCLC患者(NCT02534649)的100名NSCLC患者的肿瘤(NCT02534649),并用含有Panck,Col1a1,CD8和DDR的面板染色,然后进行图像分析(图。1 A,B)。 患者在肺腺癌(LUAD)和肺鳞状细胞癌(LUSC)中均匀分裂,以及对ICB治疗的反应者和非反应者。 根据肿瘤上皮和肿瘤基质中的CD8 T细胞密度计算连续免疫排除评分(IES)(图 1 c)。 CD8 T细胞密度(全肿瘤,肿瘤上皮和肿瘤基质)和IE在反应者与非反应器中进行了比较,并对生存率进行了评估,以鉴定ICB反应的潜在生物标志物。 在与ICB结果数据的独立肺癌队列中鉴定并测试了与IES相关的基因特征[4]。1 A,B)。患者在肺腺癌(LUAD)和肺鳞状细胞癌(LUSC)中均匀分裂,以及对ICB治疗的反应者和非反应者。根据肿瘤上皮和肿瘤基质中的CD8 T细胞密度计算连续免疫排除评分(IES)(图1 c)。CD8 T细胞密度(全肿瘤,肿瘤上皮和肿瘤基质)和IE在反应者与非反应器中进行了比较,并对生存率进行了评估,以鉴定ICB反应的潜在生物标志物。在与ICB结果数据的独立肺癌队列中鉴定并测试了与IES相关的基因特征[4]。

免疫耐受性CD8 +调节T细胞的未来前景
fi g u r e 1示意图,描绘了大鼠,小鼠和人CD8 + CD45RC低/ - treg的作用机理和标记的机理。Breg,调节B细胞;共同的,共刺激分子; DC,树突状细胞; EC,内皮细胞; IDO,吲哚胺2,3-二氧酶; Kyn,Kynurenin; MREG,调节巨噬细胞; PDC,浆细胞类动物树突状细胞; TRP,色氨酸。 弯曲的箭头表示转换或诱导。 上下箭头分别表示增加和减少Breg,调节B细胞;共同的,共刺激分子; DC,树突状细胞; EC,内皮细胞; IDO,吲哚胺2,3-二氧酶; Kyn,Kynurenin; MREG,调节巨噬细胞; PDC,浆细胞类动物树突状细胞; TRP,色氨酸。弯曲的箭头表示转换或诱导。上下箭头分别表示增加和减少

CD8 + T细胞耗尽乳腺癌的肿瘤微环境
乳腺癌(BC)是全球女性普遍的癌症,其机制尚未完全理解(1)。免疫细胞从正常乳腺组织中的免疫监视到卑诗省的进展,包括原发性和转移性阶段。卑诗省的肿瘤微环境显示出像CD4+和CD8+ GRANZYME B+细胞毒性T细胞,B细胞,巨噬细胞和树突状细胞(DCS)等免疫细胞的增加(2)。在雌激素受体(ER) - 阳性肿瘤,中性粒细胞和天然杀伤(NK)细胞中是最积极相关的,而静息的记忆T细胞和CD8+ T细胞呈负相关。相比之下,ER阴性BC显示与T调节和CD8+细胞的正相关性很强,其负相关与ER阳性病例相似(3)。值得注意的是,即使是BC早期患者,在肿瘤相关的CD8 + T细胞中也表现出精疲力尽(4),这对于消除病原体和肿瘤至关重要(5)。慢性抗原暴露和癌症中的炎症会导致CD8 + T细胞衰竭或改变分化(6),耗尽的T细胞显示肿瘤反应性和增殖


mRNA编码的持久白介素2恢复CD8 + T细胞新抗原免疫
图1:MHC I类缺乏肿瘤的免疫荒漠化和抗治疗性。(a)CT26或CT26- B2M - / - 肿瘤和免疫组织化学(IHC)T-和NK细胞浸润的纵向动力学在接种后19天对T-和NK细胞浸润进行了分析。比例尺= 50 µm。(B)接种后20天,在CT26或MC38野生型CD8 + T细胞中的PD-1表达。(c)接种19天后19天(CT26:n = 3,MC38:n = 5),在CT26或MC38野生型或B2M - / - 肿瘤组织中的IFNG表达。(d至H)用αPD-1/αCTLA4ICB组合或同种型对照(D),αPD-1,αCTLA4或IR-相关对照mab(e),GP70-nna-nna-facter(αPD-1,αCTLA4),αPD-1/αCTLA4ICB组合或同种型对照组(D),GP70-ENCORNNA-FLPX MRPX MRPX,MRNNA-FLPX,MRNNA-facter(div)(d)(d)(d至h)携带所指定的父母或b2m - / - 肿瘤变异的生存奥沙利铂/5-氟尿嘧啶(OX/5-FU)或媒介物对照(G),局部放射疗法(LRT),剂量为12 Gy或0 Gy作为对照(H)。(i)LRT(H)后9 d中的血液中的GP70抗原特异性CD8 + T细胞(n = 10)。n = 4-5每个时间点(a;左)和代表性IHC染色(a;右)。n = 8(b)。n = 3(CT26)和n = 5(MC38)(c)。这些发现表明MHC I类抗原表现的丢失,由于产生的免疫DES-

TCR捕获键非线性控制CD8合作以塑造T细胞特异性
在动态生物力学调制下,自然进化的T细胞受体(TCR)在区分非自身抗原与自我抗原方面表现出非常高的特异性。相比之下,工程设计的高级TCR通常会失去这一特殊的城市,从而与自我抗原和靶向毒性产生交叉反应。这种差异的基本机制尚不清楚。我们的研究表明,天然TCR利用机械力与其同源抗原形成最佳的捕获键。此过程依赖于机械功能的TCR - PMHC结合界面,该界面可以通过MHC和CD8中力引起的顺序构象变化,从而实现了强力增强的CD8 copector与MHC-α1α2域结合。相反,工程设计的高级tcrs与其父母TCR的同源PMHC形成了刚性,紧密结合的接口。这种刚性阻止了力诱导的构象变化,以实现最佳捕获键形成所需的构象变化。矛盾的是,这些高级的TCR可以与其父母TCR的非刺激性PMHC形成中等的捕获键,从而导致脱靶交叉反应性和降低的特异性。我们还开发了综合的力依赖性TCR - PMHC动力学功能图,能够区分功能和非功能性TCR - PMHC对并识别有毒的,交叉反应的TCR。这些发现阐明了天然TCR的特异性机械化学基础,并突出了CD8在靶向同源抗原中的关键作用。这项工作为工程TCR提供了有价值的见解,具有提高的特异性和对非自身抗原的效力,尤其是在癌症免疫疗法和传染病治疗中的应用,同时最大程度地降低了自我抗原交叉反应性的风险。