XiaoMi-AI文件搜索系统
World File Search Systemcleaved
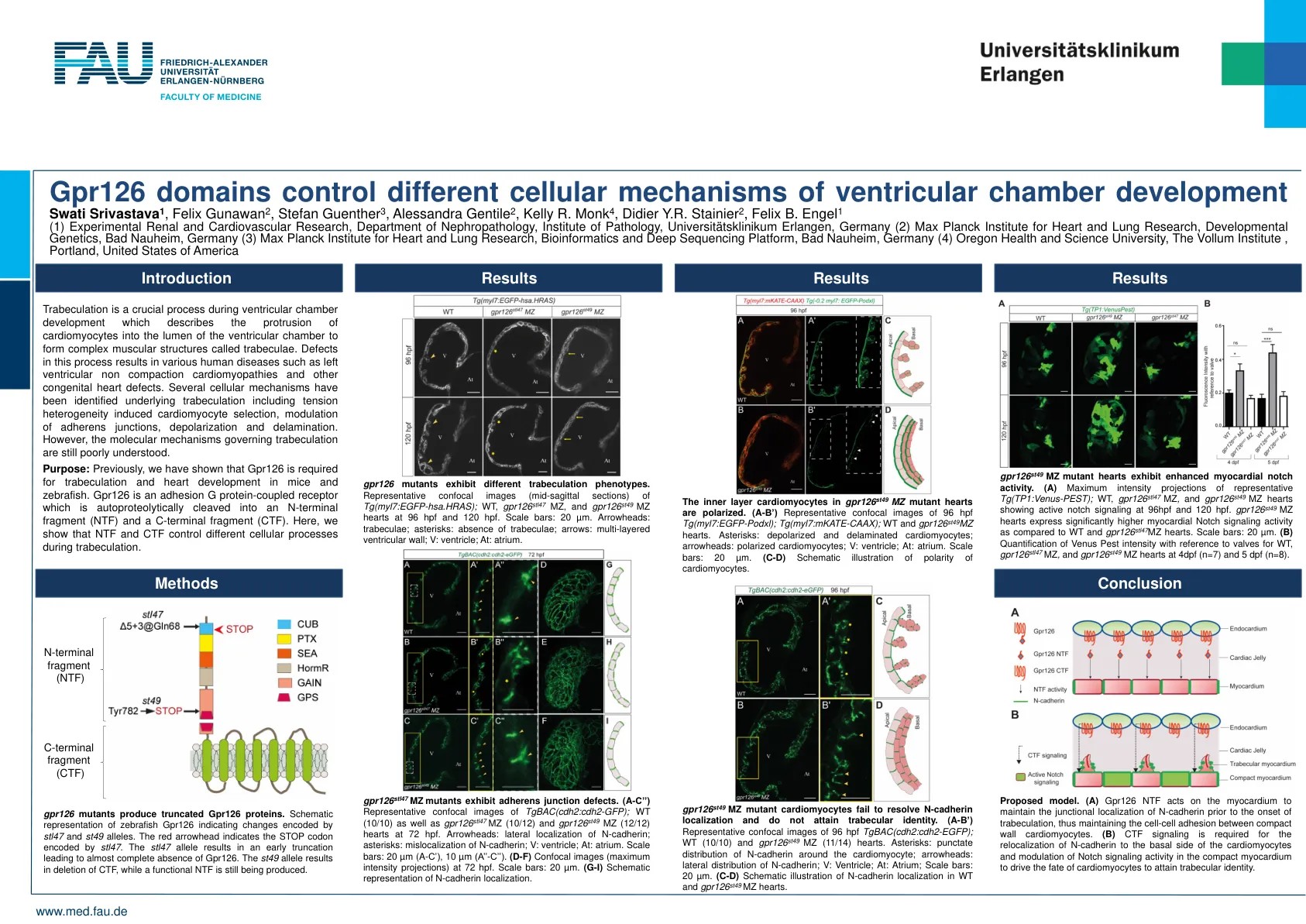
Gpr126 结构域控制心室发育的不同细胞机制 Swati Srivastava 1、Felix Gunawan 2、Stefan Guenther 3、Ale
小梁形成是心室发育过程中的一个关键过程,它描述了心肌细胞突出到心室腔内形成称为小梁的复杂肌肉结构。该过程中的缺陷会导致各种人类疾病,例如左心室非致密化性心肌病和其他先天性心脏缺陷。已经确定了小梁形成的几种细胞机制,包括张力异质性诱导的心肌细胞选择、粘附连接的调节、去极化和分层。然而,控制小梁形成的分子机制仍然不太清楚。目的:之前,我们已经证明 Gpr126 是小鼠和斑马鱼小梁形成和心脏发育所必需的。Gpr126 是一种粘附 G 蛋白偶联受体,可自蛋白酶切为 N 端片段 (NTF) 和 C 端片段 (CTF)。在这里,我们表明 NTF 和 CTF 在小梁形成过程中控制不同的细胞过程。

CAS12A的反式DNA裂解活性没有可检测到的免疫力对质粒或噬菌体
CAS12A是V-A型CRISPR-CAS RNA引导的内切酶。它在特定位点切割了dsDNA,然后在体外反式跨体内激活以非特征ssDNA的裂解。反式活性的免疫功能仍然未知。为了解决这个问题,我们在大肠杆菌中构建了一个CAS12A靶向系统,其中CAS12A裂解了高拷贝靶质粒以释放反式ssDNA裂解活性。然后,我们分析了Cas12a靶向对非目标质粒和ssDNA噬菌体的影响。结果表明,CAS12A有效地消除了目标质粒,但对噬菌体的非目标质粒或鼠疫形成的维持没有影响。此外,有助于靶质粒耗竭的两间隔crispr阵列仍然对非目标质粒或噬菌体没有可检测的影响。一起,数据表明CAS12A的反式ssDNA切割不会导致体内免疫力。

CRISPR-Cas9:基因组编辑的生物学和技术
质粒 DNA(标记为红色)被选为我们在实验室中制备的单向导 RNA 的靶序列。马丁将这些单向导 RNA 添加到纯化的 Cas9 蛋白中,并将它们与质粒 DNA 分子一起在实验室试管中孵育。为了分析该实验的结果,他在琼脂糖凝胶系统中将不同的切割 DNA 产物彼此分离,如图所示。您可以在该凝胶系统的每个泳道中看到,根据向导 RNA 与质粒 DNA 相互作用的位置,Cas9 会产生切口。通过在不同的位置切割质粒,以便同时将两个双链断裂引入质粒,我们可以将这些切割的 DNA 片段释放到凝胶系统中。如您所见,每个切割的质粒 DNA 片段根据片段的大小迁移到不同的位置。

CRISPR/CAS通过同源重组的第3章基因组编辑
在整个细胞发育中,DNA可能遭受威胁基因组完整性和细胞存活的损害。最有害的病变之一是双链DNA断裂(DSB),因为它可能导致基因组信息的丢失。DSB可能自然发生在细胞代谢期间,也可能是由外部因素触发的(Deriano; Roth,2013)。无论哪种方式,这些损坏都会通过细胞立即修复,主要是通过两种途径:非同源末端连接(NHEJ)或同源指导修复(HDR)。与通过NHEJ进行修复不同,NHEJ仅将裂解的DNA的末端连接起来(请参阅第2章),HDR途径需要存在相同或非常相似的模板,即完整的序列,以准确地修复病变的DNA(Heyer等人,2010年)。提供用于HDR中使用的模板的可能性代表了通过同源重组(HR)途径进行基因编辑的关键元素,该途径可能被利用为几种新的繁殖技术(NBT)之一。

对DNA Taphonomy的矿物学控制
fi g u r e 1云母的原子力显微镜图像显示具有不同背景离子的吸附质粒DNA的构象,DNA可见为白色(图像中的最高点)线。(a)NaCl中的云母-DNA; (b)MGCL 2中的云母-DNA; (c)NICL 2中的云母-DNA。(d)仅方解石。新鲜裂解和2分钟后。将DNA添加到表面时使用的缓冲溶液接触。图像显示步骤和蚀刻坑上的步骤边缘。(E)缓冲液中的方解石-DNA。表面显示蚀刻坑和吸附的DNA。蚀刻坑的形态取决于方解石晶体的底层正交结构,该结构的定向使钝角是向西南定向的,并且急性角针对东北定向。(f)NaCl中的方解石-DNA。(g)MGCL 2中的方解石-DNA。(H)NICL 2中的方解石-DNA。(i)在聚-L赖氨酸底物上吸附的质粒DNA。DNA是超螺旋的,并用10 mM NaCl作为背景电解质沉积。

sgRNA 的位点特异性和酶促交联可实现波长可选的光激活控制 CRISPR 基因编辑
化学交联能够快速识别 RNA-蛋白质和 RNA-核酸分子间和分子内相互作用。然而,目前尚无方法能够位点特异性和共价交联 RNA 内两个用户定义的位点。在这里,我们开发了 RNA-CLAMP,它能够位点特异性和酶促交联(夹紧)RNA 内两个选定的鸟嘌呤残基。分子内夹紧会破坏正常的 RNA 功能,而随后对交联剂进行光裂解会恢复活性。我们使用 RNA-CLAMP 通过光裂解交联剂夹紧 CRISPR-Cas9 基因编辑系统的单向导 RNA (sgRNA) 内的两个茎环,完全抑制编辑。可见光照射会裂解交联剂并以高时空分辨率恢复基因编辑。设计两种对不同波长的光有响应的光裂解接头,可以在哺乳动物细胞中实现基因编辑的多路复用光激活。这种光激活的 CRISPR-Cas9 基因编辑平台受益于无法检测的背景活动,提供激活波长的选择,并具有多路复用功能。

一种优化的定量蛋白质组学方法建立了细胞类型的小鼠脑分泌
要了解细胞如何在神经系统中进行通信,必须定义其分泌组,这对于原代细胞的挑战很大,因为需要大细胞数。在这里,我们通过开发“用点击糖的高性能分泌蛋白富集”(HISPECS)方法来小型分泌组分析。为了证明其广泛的效用,HISPEC用于确定脑切片对LPS诱导的神经炎症的分泌反应,并使用原代星形胶质细胞,微神经胶质,神经元和寡聚细胞来建立细胞类型分辨的小鼠脑秘密资源。该资源允许映射CSF蛋白的细胞起源,并揭示出意外的体外和体内分泌的蛋白质出乎意料的是蛋白水解裂解的膜蛋白extodomain。两个例子是神经分泌的Adam 22和CD 200,我们将其确定为阿尔茨海默氏症连接蛋白酶贝丝的底物1。hispecs和脑部分泌资源可以被广泛利用,以系统地研究蛋白质分泌和脑功能,并鉴定CNS疾病的细胞类型特异性生物标志物。
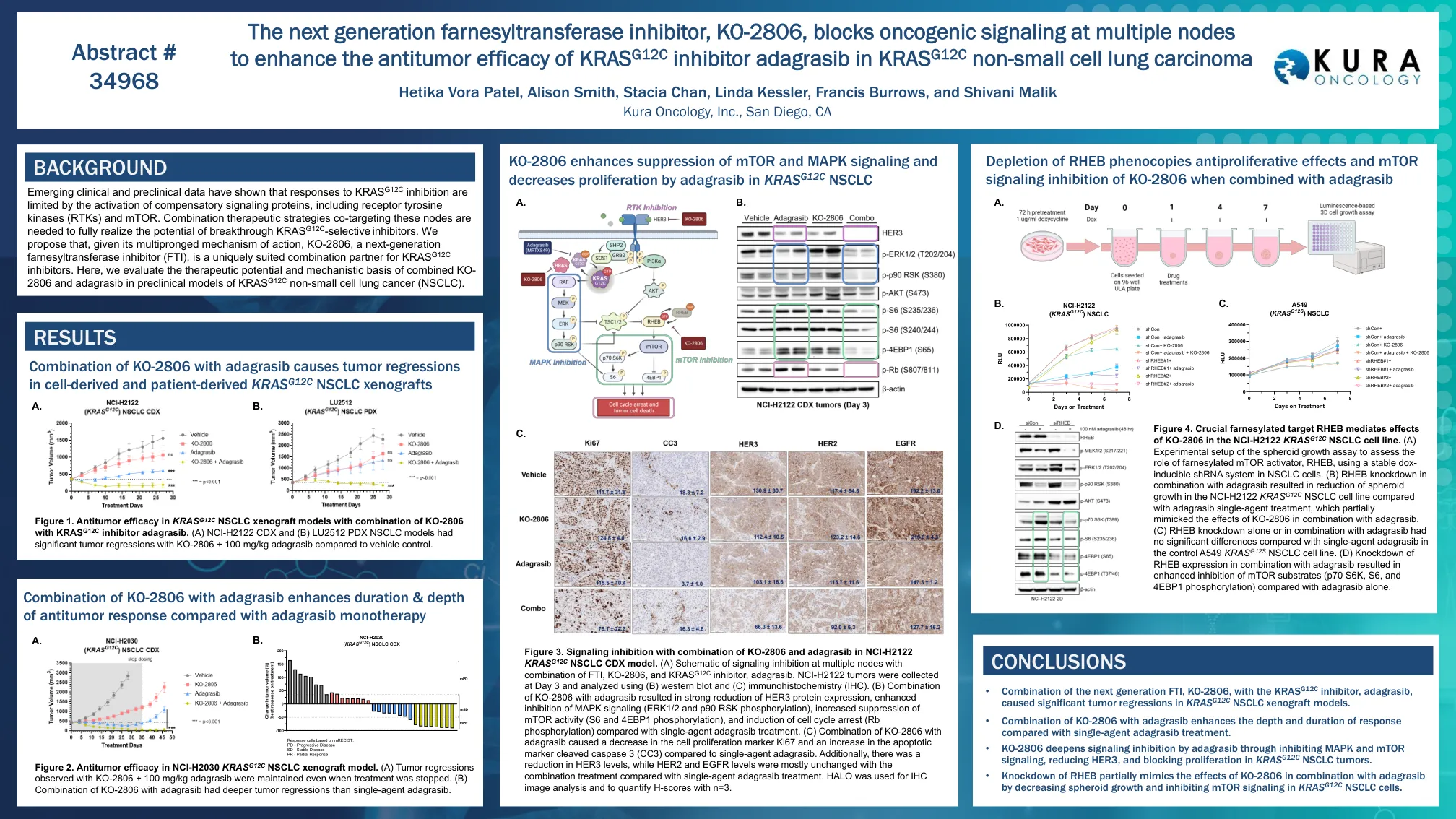
下一代Farnesylsylansferase抑制剂KO-2806,阻止多个节点的致癌信号传导,以增强KRAS G12的抗肿瘤功效
图3。NCI-H2122 KRAS G12C NSCLC CDX模型中KO-2806和Adagrasib结合使用的信号传导抑制。(a)在多个节点上抑制信号传导的示意图与FTI,KO-2806和KRAS G12C抑制剂Adagrasib的组合。NCI-H2122肿瘤,并使用(b)Western印迹和(c)免疫组织化学(IHC)分析。(b)KO-2806与AdagrasiB的组合导致HER3蛋白表达的大幅度降低,增强了对MAPK信号传导(ERK1/2和P90 RSK磷酸化)的抑制作用,增加了MTOR活性的抑制(S6和4EBP1磷酸化),以及对单个Cell Cycer Cyprant(RB Pranscratib)的抑制。(c)KO-2806与Adagrasib的组合导致细胞增殖标记Ki67的降低,而与单药adagrasib相比,凋亡标记裂解的caspase 3(CC3)的增加。此外,与单药Adagrasib治疗相比,HER3水平降低了,而HER2和EGFR水平的组合处理大多是没有变化的。halo用于IHC图像分析,并用n = 3量化H得分。

cnrs在工作人员的火下
1973),他指出,当将金属和半导体放置在亲密接触中时,它们的表面都将被改变,尤其是通过电子从金属穿透到半导体表面的能力,通过隧穿进入禁止的能量隙。这种效果改变了电荷分布,因此可以以质量上很大的结果来解释界面处的电场。逐渐清楚地表明,定量的效果不足以解释地表状态的数量或空间分布。Thanailakis的结果(J.Phys。C. 8,655; 1975年),是确定这一点最重要的之一。我们现在提出了最新建议,并得到了斯坦福大学Spicer及其小组的广泛和令人印象深刻的实验数据的支持。最新纸(Spicer等人物理。修订版Lett。 44,420; 1980)阐明了他们在许多复合半导体中提出的屏障形成的机制。 这些材料在禁止能量隙中没有内在的表面状态。 也就是说,在这些材料的分裂表面上,Fermi能量被发现Lett。44,420; 1980)阐明了他们在许多复合半导体中提出的屏障形成的机制。这些材料在禁止能量隙中没有内在的表面状态。也就是说,在这些材料的分裂表面上,Fermi能量被发现

DNA修复的全基因组分析标识更高 -
(a)实验设置和集成的概述。(b)1p染色体上的信号。左:在 +DSB条件下的单细胞热图(RPKM),其顶部为 +DSB(有色)和–DSB(灰色)条件的单细胞聚集体。右:带有覆盖MSR调用的单细胞线图。asisi主题用黑线注释,红色三角形表示经常裂解(或“顶部”)位点。(c)所有修复频率≥10%的ASISI位点的条形图,每个位点的修复频率(目标蛋白质和方法)颜色为颜色。通过增加绝对修复频率(即任何数据集中的最高频率)来订购(在X轴上)。每个站点,通过增加每个数据集的维修频率(前后;即未堆叠栏)来排序条。底部水平条表示先前的(缺乏)注释作为顶部位点。(d)一个代表性核的共聚焦图像显示DAPI,RAD51 DAMID M6 A-TRACER和内源性γH2AX免疫荧光染色。(e)信号共定位(Manders的A和B每个核)的定量,n = 33核。