XiaoMi-AI文件搜索系统
World File Search Systemcod

印度尼西亚绿色分类法的下一阶段:支持脱碳的过渡融资
a。到2040年的煤炭阶段; 2022年12月31日之后建造的植物将无资格; b。运营限制在商业运营日期(COD)c。合格的植物必须:展示最佳的班级技术(负担得起,易于访问,可靠),这些技术由国际公认的机构独立验证,因为它在预期的寿命(ADB ETM或JETP下的植物符合这些标准下)具有积极的排放能力(符合这些标准)
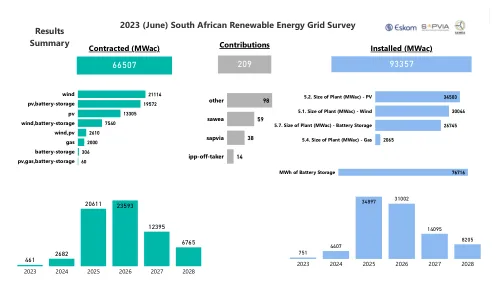
2023(6月)南非可再生能源网格调查
批准了环境批准/决策记录(ROD)(愿意提供申请参考号的贡献者)。现场测量活动和可行性工作已经完成。ppa签名或接近签名/或项目将准备竞标到最近的Reippp回合。如果Eskom或市政当局立即获得网格连接,则此类别的项目将能够在3年内到达COD。对于嵌入式生成项目,该项目将在3年内随时连接到嵌入式网络。
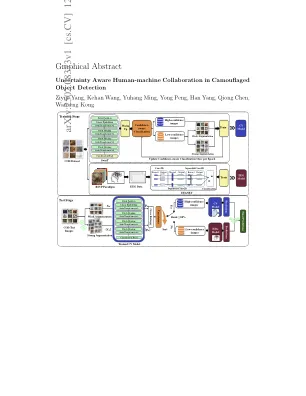
不确定性意识到伪装对象检测中的人机合作
伪装的对象检测(COD)是识别在其环境中识别对象的任务,由于其广泛的实际应用范围很快。开发值得信赖的COD系统的关键步骤是对不确定性的估计和有效利用。在这项工作中,我们提出了一个人机协作框架,用于对伪装物体的存在进行分类,利用计算机视觉模型(CV)模型的互补优势和无创的脑部计算机界面(BCIS)。我们的方法引入了一个多视障碍,以估计简历模型预测中的不明显,利用这种不确定性在培训过程中提高效率,并通过基于RSVP的BCIS在测试过程中为人类评估提供了低信任案例,以实现更可靠的决策。我们在迷彩数据集中评估了框架,与现有方法相比,平衡准确性(BA)的平均平均提高为4.56%,F1得分的平均提高为4.56%。对于表现最佳的细节,BA的改善达到7.6%,F1分数为6.66%。对培训过程的分析表明,我们的信心措施和精度之间存在很强的相关性,而消融研究证实了拟议的培训政策和人机合作的有效性

利用可再生能源的规则
(i)任何建立以煤炭/褐煤为基础的火力发电站的发电公司,若项目的商业运营日期(COD)在 2023 年 4 月 1 日或之后,则须建立可再生能源发电能力(以兆瓦为单位),即可再生能源发电义务(RGO)至少为以煤炭/褐煤为基础的火力发电站容量(以兆瓦为单位)的百分之四十 (40%),或采购和供应相当于该容量的可再生能源。

估计土狼饮食和对威胁的捕食
随着由于气候变化和人类对景观的修改,世界继续变化,一些物种已受到威胁或灭绝,而另一些物种在这些新条件下蓬勃发展。土狼(Canis Latrans)自1900年代初以来就扩大了整个北美的范围,并于1970年代到达马萨诸塞州科德角。位于鳕鱼外角的鳕鱼角国家海滨是一个受保护区域,其中包含两个威胁性shore鸟物种的重要嵌套栖息地:最小的tern(胸骨antillarum)和管道plover(Charadrius Melodus)。人类的景观修饰,捕食和其他因素导致两种栖息地物种的下降和范围下降。土狼是一种机会主义的杂食动物,消耗了其环境中最容易获得的东西,包括潜在的shore鸟。然而,该生态系统中土狼捕食的毛鸟捕食程度仍然未知。为了了解土狼对受保护的海鸟的潜在影响,我们使用DNA元法编码分析了土狼饮食。这项研究的目标是(i)评估受威胁的海鸟对土狼饮食的存在和贡献,以及(ii)检查土狼饮食中的季节性和基于性别的变化。,我们在2022年秋天(景观上不存在海岸鸟)和2023年夏季(当时在景观上存在海岸鸟)并使用metabarcododing估算饮食饮食组成。我们首先将scat样品构成,以确认物种并基因分类,以识别性别和个体。我们使用样品子集的元法编码来识别shore鸟和其他脊椎动物的存在。总共我们在秋季收集了215个SCAT样本,在夏季收集了213个SCAT样本,分别确定了57个和55个独特的人(两个季节中检测到的21个人)。我们选择了从尽可能多的不同个体中选择的样品来进行元编码,从而从每个独特鉴定的土狼中提供至少一个样本。

q q q - ASSIST 快速搜索
折射率,最小1.3630 1.3ss0 4.7。粘度,硅酸盐 ASTM D445-74 4.7.2 最小值 5 “C 20 10 最小值 25'C 2 2 氢离子浓度(PI) 7.0 至 8.5 7.0108.5 4.7.3 扩散系数,最小值 3 3 4.7.4 成形性 泡沫膨胀,最小值 5.0 S.o NFTA STD 412 4.7.5 泡沫 25% 排水时间,最小值,最小值 2.5 2.5 NFPA STD 412 4.7.5 腐蚀性 常规 冷轧,低碳钢 SICCI(UNS G 10-1OO),高强度,最大 I .5 1.5 ASTM E527 4.7.7 铜镍合金(90-10)(UNS C70600),微小损伤,最大值 I.0 1.0 ASTM S-S27 4.7.7 N,ckel-ppcr (70-30) (UNS N04400),微小损伤,最大值 I.0 I.0 ASTM E-527 4.7,7 青铜 (UNS C90500),毫克,最大值 100 100 ASTM ES27 4.7,7 耐腐蚀,MnSb (CRES) 雪橇,(UNS S304fXJ) 无凹坑 无损伤 4.7.7 总卤化物-p/m,最大值 210 ASTM D1821 4.7.8 干化学耐久性,燃烧耐受时间,秒,最小值 360 360 4.7.9 环境影响:毒性,LC50 m#L,最小 SW moo 4.1.12.1 COD,mg/L,最大 1000K 500K 4.7.12.2 ~20 最小 .65 .65 COD 4,7.123

热预处理对通过水解 - 酸发生从棕榈油磨坊流出物(POME)产生生物氢的影响
摘要:作为世界上最大的棕榈油生产商之一,印度尼西亚具有利用棕榈油厂废水(POME)的巨大潜力,以生产氢作为有希望的能源。这项研究研究了热预处理对从水解到水解 - 累积发生的生物氢产生效率的影响。在与牛粪结合之前,在各种温度(50、75、100、125和150°C)的各种温度(50、75、100、125和150°C)上进行了预处理。将组合在35°C的生物反应器中发酵48小时。每四个小时,使用GC-TCD监测氢气的产生,并在反应前后对底物的化学氧需求(COD)进行研究,以确定预处理的效率。研究发现,将材料预热至100°C可产生最佳效果,氢含量为36.5%,COD去除效率为22.74%。最高的氢产率为每升氢氢的0.264升,这是理论最大值的8.79%。当温度超过100°C时,由于形成了顽固的物质,氢产生降低。这些发现强调,正确的热预处理可以极大地增强POME的生物氢产生,从而提供一种可持续的方法来管理废物并产生替代能源。

奥尔良镇 - 十年住房计划执行摘要
该计划利用来自许多来源的数据,包括 2010 年和 2020 年每十年一次的人口普查、2021 年美国社区调查、当地建筑许可数据、当地评估员数据、当地分区条例、住房和宜居社区执行办公室、综合住房可负担性战略 (CHAS)、当地 MLS 数据、Rentometer、奥尔良住房管理局、马萨诸塞州小学和中学教育部 (DESE)、HUD、马萨诸塞州住房合作伙伴的 DataTown、该镇网站、大波士顿住房报告卡 2022、麻省大学多纳休研究所数据、科德角委员会奥尔良数据、UMDI 的 2023 年科德角住房需求分析、马萨诸塞州经济研究部数据、之前的城镇计划(报告中引用)以及工作人员和委员会的知识和数据。美国人口普查通过询问十个问题来统计美国的每一位居民,而 ACS 则根据人口样本提供估计值以获取更详细的信息。需要注意的是,ACS 估算值包含误差幅度 (MOE),因为它们基于样本,而非完整计数。数据收集和分析是在 2023 年夏季进行的,本报告反映了当时的最新数据,仅包括可用的 2020 年十年一次人口普查信息。
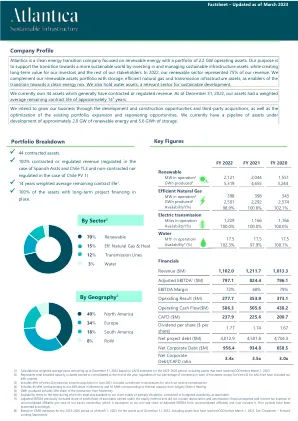
概况介绍 - Atlantica 可持续基础设施
(1) 根据 CAFD 对 2023-2026 年期间的估计,计算为截至 2022 年 12 月 31 日的加权平均剩余年限,包括 2023 年 3 月 1 日之前已达到 COD 的资产。 (2) 代表年末拥有或合并的资产的总装机容量,无论我们在每项资产中的所有权百分比是多少,但 Vento II 除外,我们已将 49% 的权益计入其中。 (3) 包括自 2021 年 6 月收购 Vento II 以来的 49% 产量。包括我们获得补偿的风电资产削减 (4) 包括 43 MW,相当于我们在蒙特雷的 30% 份额,以及 55 MWt,相当于卡尔加里区域供热的热容量。 (5) 生产的 GWh 包括蒙特雷生产的 30% 份额。 (6) 可用性是指资产全部或部分可供客户使用的时间除以合同或预算可用性(如适用)。 (7) 调整后的 EBITDA 之前不包括按权益法核算的联营公司利润/(亏损)份额,也不包括非合并联营公司的折旧和摊销、财务费用和所得税费用(按我们的股权比例计算)(相当于我们按比例从非合并联营公司获得的调整后 EBITDA),现在包括了这些费用。前期已相应列示。 (8) 根据 CAFD 对截至 2023 年 3 月 1 日的 2023-2026 年期间资产的估计,包括 2023 年 3 月 1 日之前已达到 COD 的资产。请参阅“免责声明 - 前瞻性陈述”。

利用 - 吸收 - 氧化过程和净过滤 -
粮食生产的快速工业化已经显着影响了各个部门产生的废水的质量和数量。由于其废水废水引起的环境问题引起了环境问题的一个这样的行业是食品行业,尤其是酱油生产行业。酱油是许多亚洲美食中的主食调味品,其生产涉及复杂的发酵过程,通常导致废水高度颜色,化学化,化学复杂且充满了有机污染物。浪费酱油是酱油制造的副产品,以其高化的化学氧需求(鳕鱼),高水平的有机化合物和浓烈的色彩而闻名,所有这些都会有助于环境污染,如果不正确地管理以应对处理浪费酱油的兴趣和越来越多的兴趣,则对先进的氧化物进行了越来越多的兴趣,并且利用了先进的氧化能力,即利用先进的水平(一定的水)。 (NF),以提高废水质量。AOP包括诸如臭氧化,紫外线(UV)轻处理和芬顿试剂之类的过程,在分解复杂的有机污染物,减少鳕鱼和脱色废水方面非常有效。另一方面,纳米过滤是一种基于膜的分离过程,能够从水中去除溶解的盐,有机分子和颗粒物,使其在废水处理的背景下成为有价值的技术[1]。