XiaoMi-AI文件搜索系统
World File Search Systemcortical

皮质视觉障碍 (CVI) 适应材料 - SPEVI
• 由于 CVI 的性质,许多患有该病的人可能无法获得标准材料 • 基于眼部损伤的干预和调整可能无法为患有 CVI 的人提供适当的支持 • 根据评估提供改编的材料非常重要

急性疼痛期间诱发的震荡皮质活动
Daniel Ciampi de Andrade 神经可塑性和疼痛中心 (CNAP) 奥尔堡大学医学院健康科学与技术系 Selma Lagerløfs Vej 249 9260 Gistrup 丹麦 电子邮件:dca@hst.aau.dk

Cortical networks of dynamic scene category representation in the human brain
视觉神经科学的主要目的是阐明人脑在自然场景中如何代表各种信息。对场景感知的行为研究表明,人类将场景归类为更有效地处理视觉场景中的大量信息(Greene&Oliva,2009; Konkle,Konkle,Brady,Alvarez,&Oliva,&Oliva,&Oliva,2010; Rousselet,Joubert,&Fabre-Thorpe,2005年)。因此,场景类别上的信息可能在皮质上表示。与这一概念一致,以前的神经影像学研究表明,视觉场景的类别可以在基于血液氧气水平(大胆的)响应(大胆的)响应中的有限数量的基本类别(例如,海滩,森林,山脉)之间进行分类。 OPA), object-selective lateral occipital complex (LO), and anterior visual cortex ( Epstein & Morgan, 2012 ; Jung, Larsen, & Walther, 2018 ; Walther, Caddigan, Fei e Fei, & Beck, 2009 ; Walther, Chai, Caddigan, Beck, & Fei e Fei, 2011 ).这些研究中的一种常见方法是将视觉场景定义为几个非重叠类别。然而,自然场景可能显示出不同程度的统计相关性,并且在几个不同的类别下可能会表征现实世界的场景。此外,由于这些研究使用了静态场景,因此它们没有必要的工具来证明人类大脑中的动态场景类别是如何代表的。Stansbury等。要检查自然场景类别的统计数据,最近的一项研究(Stansbury,Naselaris和Gallant,2013年)使用了数据驱动的算法来采购广泛的场景,其中还考虑了这些类别之间的潜在相似性。在这种方法中,将每个场景类别定义为在自然场景中出现的大量组成对象的存在概率列表。一旦算法学习一组类别,就可以根据场景中的对象来推断给定场景属于每个学到的类别的可能性。据报道,与基于一些经常出现在提出的自然图像中经常出现的诊断对象的存在相比,与替代模型相比,经典面部和场景选择区域中单素大胆响应的预测得到了改进的预测(Stansbury et al。,2013年)。此结果提高了对象共发生统计构成场景的基础的可能性。通过其组成对象定义了静态场景的类别,并着重于经典场景选择区域中的类别响应,例如许多先前关于场景代表的研究(Epstein&Morgan,2012; Jung et al。,2018; Walther等,2009,2011,2011; Jung et al。,2018; Jung et al。然而,最近的几项研究表明,大部分前视觉皮层可能是通过对视觉场景中的动作进行差异调整来组织的(Tarhan&Konkle,2020;CáUkur,Huth,Huth,Nishimoto和Gallant,2016年)。实际上,现实世界的场景包含对象和动作之间的动态影响(Greene,Baldassano,Esteva,Beck,Beck,&Fei E Fei,

小鼠头部撞击过程中双脑皮质钙成像...
我们提出了一种经济高效、体积小巧、基于开源 Raspberry Pi 的宽视野成像系统。紧凑的特性使该系统可用于近距离双脑皮质中尺度功能成像,以同时观察两只头部固定的动物在分阶段的社交接触式互动中的活动。我们提供了轨道系统的所有原理图、代码和协议,其中头部固定的小鼠被带到一定距离,每只小鼠的大触须都会接触。在社交接触期之前、期间和之后,同时记录了两只小鼠的皮质神经元功能信号 (GCaMP6s;遗传编码的 Ca 2 1 传感器)。当小鼠在一起时,我们观察到了相互拂动和跨小鼠相关皮质活动的发作。在试验打乱的小鼠对中未观察到相关性,这表明相关活动特定于个体互动。在小鼠在一起(最密切接触)期间观察到与拂动相关的皮质信号。社会刺激呈现的影响延伸到与相互接触相关的区域之外,并对皮质活动产生整体同步效应。

皮层手图的延伸性,遵循指导神经阻滞
猴子中的电生理研究表明,手指截肢会触发局部重塑,内部的原发性体感皮质(S1)。人类神经影像学研究表明,即使在截肢后数十年,也表明了缺失手指的持续代表。在这里,我们探讨了这种明显的矛盾是否源于低估手部图中手指的分布的外围和中心表示。使用药理学单指神经阻滞和7-Tesla神经影像学,我们首先复制了局部S1重新映射的先前帐户(电生理和其他)。局部阻塞还触发了整个手部区域的活动的活动变化。使用利用指定代表重叠的方法,我们还表明,尽管输入损失,但被阻塞的手指表示仍然持久。计算建模表明,局部稳定性和全局重组都是由地形图基础的分布处理以及稳态机制结合的。我们的发现揭示了复杂的指代代表性特征,这些特征在大脑(RE)组织,超越(RE)映射中起着关键作用。

psilocybin增强恐惧灭绝与皮质合奏的双向调制有关
5-羟色胺2受体(5HT2R)激动剂psilocybin在神经术 - 跨科学疾病中表现出快速而持续的治疗疗效,这些疾病的特征是认知僵化。然而,尚未表征psilocybin对行为灵活性持续变化的神经活动模式的影响。测试了psilocybin通过改变皮质神经合奏中活性增强行为柔韧性的假设,我们在为期五天的痕量恐惧学习和灭绝测定中进行了延误皮质中的纵向单细胞成像。一剂psilo-cybin引起的恐惧学习和灭绝日之间的集合周转,同时相反地调节了恐惧和灭绝活性神经元中的活动。急性抑制恐惧活性神经元和延迟灭绝活性神经元的募集是预测psilocybin增强恐惧灭绝的。一个计算模型表明,psilocybin对恐惧活性神经元的急性抑制足以解释其几天后其神经和行为效应。这些结果与我们的假设保持一致,并引入了一种新的机制,涉及回合膜皮质中恐惧活性种群的抑制。

通过人类生命跨度的功能连接
标题:青少年和年轻人的皮质旋转与睡眠之间的关联。作者:JoãoPauloLima Santos 1,医学博士;丽贝卡·海斯(Rebecca Hayes),博士1,彼得·L·弗朗森(Peter L. Franzen),博士1,蒂娜·R·戈德斯坦(Tina R. Dana L. McMakin,博士6,Neal D. Ryan,MD 1,Jennifer S. Silk,PhD 2,Maria Jalbrzikowski 7,8*,博士学位; Adriane M Soehner 1*,博士学位。隶属关系:1。美国匹兹堡大学精神病学系,美国宾夕法尼亚州匹兹堡; 2。宾夕法尼亚州匹兹堡匹兹堡大学心理学系; 3。匹兹堡大学临床与转化科学研究所,
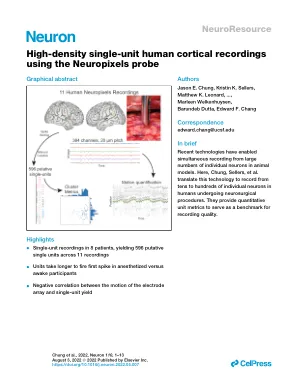
使用 Neuropixels 探针进行高密度单元人类皮质记录
动作电位是神经计算的基本单位。尽管在动物模型中记录大量单个神经元方面已经取得了重大进展,但由于临床限制和电极可靠性,这些方法在人类身上的转化受到限制。在这里,我们介绍了一种可靠的方法,使用 Neuropixels 探针在人类术中记录数十个神经元,可同时记录多达 100 个单个单元。大多数单个单元在到达目标深度后 1 分钟内处于活动状态。电极阵列的运动与产量呈很强的负相关性,这表明进一步提高探针效用面临着重大挑战和机遇。在大多数记录中,时间上活动相近的细胞对在空间上也更接近,展示了解决复杂皮质动态的能力。总之,这种方法可以访问人类新皮质深度上的群体单个单元活动,而这种规模以前只能在动物模型中访问。

语言时间尺度的皮质表征在阅读和听力之间共享
自然语言理解被认为涉及将低级感官表征整合到越来越高级的表征层次中。为了研究人类大脑是否使用相同的通路来整合书面语言和口语,我们测试了这种层次结构的大脑表征是否在书面语言理解和口语理解之间共享。语言层次结构的不同级别在操作上被定义为时间尺度,其中每个时间尺度指的是语言刺激的频谱成分,这些成分在一定数量的单词中变化。体素编码模型用于确定不同语言时间尺度在大脑皮层中的表征方式,针对每种模态分别进行。编码模型是使用 fMRI BOLD 数据估计的,这些数据是在参与者阅读和聆听每种模态中的同一组自然语言叙述时记录的( Huth 等人,2016 年;Deniz 等人,2019 年)。这些模型揭示了两种模态之间皮层表面语言时间尺度表征的拓扑组织高度相似。我们的结果表明,大脑对语言时间尺度的表征很大程度上与刺激方式无关。

DNA复制叉速度在皮质神经发生中充当发动机
a,示意图,显示了MCMBP介导的组装,并将MCM3-7导出到核中,该核能形成新生的MCM,用MCM2作为恢复前复合物,并调节DNA复制叉速度。nls表示核定位信号。b,从顶端到基础位置的MCMBP的时空表达,从E12.5到E15.5。c,蛋白质印迹分析显示了皮质发育产前和产后阶段的MCMCBP表达模式。d,在P3处的CKO小鼠和同窝对照的代表性图像。红色星星指示CKO鼠标。e,(左图)MCMBP +/ +的背视图; EMX1-CRE和MCMBP FL/FL; EMX1-CRE(CKO)P4大脑。(右图)与同窝对照(CTRL)相比,CKO中的皮质区域显着降低。(平均,两尾未配对的t检验,ctrl:n = 7,cko:n = 5)。f,(左图)MCMBP +/ +和CKO P4脑的DAPI染色冠状切片。与同窝对照(CTRL)相比,CKO的皮质板厚度显着降低了皮质板厚度。(平均,两尾未配对的t检验,ctrl:n = 7,cko:n = 5)。g,MCMBP +/ +的P4脑中的层标记物BRN2,TBR1,LHX2和TLE4的免疫染色; EMX1-CRE和CKO。h,与同窝对照组(CTRL)相比,CKO的上层神经元显着降低。(均值,两尾未配对的t检验,BRN2,TBR1,CTRL:n = 8,cko:n = 5,lhx2,tle4,ctrl:n = 4,cko:cko:n = 4)。i,蛋白质印迹分析显示了E15.5,E16.5和P4 Cortex中MCMCBP表达的下调。(平均,两尾未配对的t检验,ctrl:n = 3,cko:n = 3)。J,MCMBP +/ +中的顶祖细胞标记物SOX2和中间祖细胞标记的免疫染色; EMX1-CRE和CKO从E12.5到E16.5。K,SOX2+细胞数分析表明,在E12.5处CTRL和CKO之间没有差异。但是,由于E13.5,Sox2+细胞显着降低并持续到E16.5。(mean, two-tailed unpaired t-test, E12.5, ctrl: n=5, cKO: n=4, E13.5, ctrl: n=4, cKO: n=3, E14.5, ctrl: n=5, cKO: n=5, E15.5, ctrl: n=6, cKO: n=4, E16.5, ctrl: n=6, CKO:n = 4)。l,EOMES+细胞数分析表明,在E12.5和E13.5处CTRL和CKO之间没有差异。但是,Eomes+细胞从E14.5显着降低到E16.5。(mean, two-tailed unpaired t-test, E12.5, ctrl: n=3, cKO: n=3, E13.5, ctrl: n=4, cKO: n=4, E14.5, ctrl: n=4, cKO: n=4, E15.5, ctrl: n=4, cKO: n=3, E16.5, ctrl: n=4, CKO:n = 3)。