XiaoMi-AI文件搜索系统
World File Search SystemeGFP

利用 CRISPR/Cas9 介导的靶向整合生成 Y 染色体中含有 EGFP 基因的转基因克隆水牛胚胎
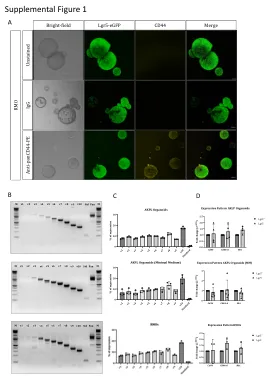
性别控制技术在家畜生产中具有重要意义,尤其对于快速繁殖的水牛(bubalus bubalis)具有重要意义,本研究以水牛为研究模型。我们已证实整合到小鼠Y染色体上的荧光蛋白可用于小鼠植入前胚胎的性别鉴定。首先,我们优化了增强型绿色荧光蛋白(eGFP)和mCherry外源基因在Neuro-2a细胞、小鼠胚胎干细胞、小鼠胚胎细胞(NIH3T3)、水牛胎儿成纤维细胞(BFF)中的靶向整合效率。结果表明,靶标两侧同源臂长度为800 bp比300 bp或300 bp/800 bp更有效。当细胞补充了 pifithrin-µ(一种抑制 p53 与线粒体结合的小分子)时,BFF 细胞中同源定向修复 (HDR) 介导的敲入也得到了显著改善。250 V 的三个脉冲在 BFF 细胞中产生最有效的电穿孔,并且发现 1.5 µ g/mL 嘌呤霉素是筛选的最佳浓度。此外,利用 CRISPR/Cas9 介导的基因编辑结合体细胞核移植 (SCNT) 技术成功生成了 Y-Chr-eGFP 转基因 BFF 细胞和克隆水牛胚胎。在第 6-8 代时,Y-Chr-eGFP 转基因 BFF 细胞的生长率和细胞增殖率明显低于非转基因 BFF 细胞;甲基化相关基因 DNMT1 和 DNMT3a 的表达水平相似;然而,与非转基因细胞相比,Y-Chr-eGFP 转基因 BFF 细胞中乙酰化相关基因 HDAC1 、 HDAC2 和 HDAC3 的表达水平显著较高(p < 0.05)。Y-Chr-eGFP 转基因 BFF 被用作 SCNT 的供体,结果表明 eGFP 报告基因适用于胚胎性别的可视化。克隆水牛胚胎的囊胚率相似;然而,与对照相比,转基因克隆胚胎的卵裂率明显较低。总之,我们优化了产生转基因 BFF 细胞的方案,并使用这些细胞作为供体成功产生了 Y-Chr-eGFP 转基因胚胎。

优化的 HDR 介导的 K-荧光蛋白敲入...
图 2:在 K-562 细胞中通过电穿孔优化 HDR 介导的报告基因敲入。A. LMNA -EGFP 供体质粒单独电穿孔,浓度增加,显示非特异性 EGFP 表达量较低。Cas9 蛋白:crRNA:tracrRNA 电穿孔,LMNA -EGFP 供体质粒浓度增加,显示 EGFP 表达相关增加。B. 放大倍数增加后,HDR 介导的敲入样本中 EGFP 表达的定位与 LMNA 的预期一致,位于细胞核中,并与 Hoechst 染色共定位。C. HDR 介导的敲入细胞群的流式细胞术分析显示,随着 DNA 供体质粒数量的增加,EGFP 表达增加高达 32%。单独 DNA 供体质粒对照的相应分析显示所有剂量的 EGFP 表达均低于 0.5%(未显示数据)。

新型转基因斑马鱼系用于研究 CHRNA3‐...
摘要 乙酰胆碱 (ACh) 是周围神经系统 (PNS) 和中枢神经系统 (CNS) 的重要神经递质,它通过烟碱型乙酰胆碱受体 (nAChR) 和毒蕈碱型乙酰胆碱受体 (mAChR) 发出信号。在这里,我们探讨了三个 nAChR 亚基 chrna3 、 chrnb4 和 chrna5 的表达模式,它们位于进化保守的簇中。在多种脊椎动物中,这种紧密的基因组定位可能表明共同功能和/或共同表达。通过新型转基因斑马鱼系,我们观察到 PNS 和 CNS 内广泛表达。在 PNS 中,我们观察到 chrna3 tdTomato 、chrnb4 eGFP 和 chrna5 tdTomato 在肠道神经系统中的表达; chrna5 tdTomato 和 chrnb4 eGFP 位于侧线的感觉神经节中;而 chrnb4 eGFP 位于耳朵中。在中枢神经系统中,chrnb4 eGFP 和 chrna5 tdTomato 的表达出现在视网膜中,这三种基因均在大脑的不同区域表达,其中一部分 chrna3 tdTomato 和 chrnb4 eGFP 细胞被发现是投射到侧线的抑制性传出神经元。在脊髓内,我们在运动网络内识别出表达 chrna3 tdTomato、chrnb4 eGFP 和 chrna5 tdTomato 的不同神经元群,包括表达 dmrt3a 的中间神经元和表达 mnx1 的运动神经元。值得注意的是,每个半节段的三到四个初级运动神经元均被 chrna3 tdTomato 和 chrnb4 eGFP 标记。有趣的是,我们在每个半节段中发现了一个 sl 型次级运动神经元,该神经元强烈表达 chrna5 tdTomato 并同时表达 chrnb4 eGFP。这些转基因系为 nAChRs 在运动网络中的潜在作用提供了见解,并为探索它们在整个神经系统一系列组织中尼古丁暴露和成瘾的作用开辟了途径。

开发用于高内涵筛选的 CRISPR/Cas9 介导荧光报告基因人类多能干细胞系
摘要:应用 CRISPR/Cas9 系统将荧光蛋白敲入人类多能干细胞 (hPSC) 中的内源性目的基因,有可能促进基于 hPSC 的疾病建模、药物筛选和移植疗法优化。为了评估荧光报告 hPSC 系用于高内涵筛选方法的能力,我们将 EGFP 靶向内源性 OCT4 基因座。产生的 hPSC–OCT4–EGFP 系表达与多能性标记物一致的 EGFP,并且可以适应多孔格式以进行高内涵筛选 (HCS) 活动。然而,在长期培养后,hPSC 暂时失去了 EGFP 表达。或者,通过将 EGFP 敲入 AAVS1 基因座,我们建立了稳定且一致的 EGFP 表达 hPSC–AAVS1–EGFP 系,该系在体外造血和神经分化期间保持 EGFP 表达。因此,hPSC–AAVS1–EGFP 衍生的感觉神经元可适应高内涵筛选平台,该平台可应用于高通量小分子筛选和药物发现活动。我们的观察结果与最近的发现一致,表明在 OCT4 基因座进行 CRISPR/Cas9 基因组编辑后会出现高频率的靶向复杂性。相反,我们证明 AAVS1 基因座是 hPSC 中的安全基因组位置,具有高基因表达,不会影响 hPSC 质量和分化。我们的研究结果表明,应应用 CRISPR/Cas9 整合的 AAVS1 系统来生成稳定的报告 hPSC 系以用于长期 HCS 方法,并且它们强调了仔细评估和选择应用的报告细胞系以用于 HCS 目的的重要性。

神经嵴衍生细胞在伤口愈合过程中具有向角质形成细胞分化的潜能
神经嵴衍生细胞(NCDC)在胎儿期以神经嵴细胞的形式存在,并分化为腭细胞,也存在于成人腭组织中,但其作用尚不明确。本研究用EGFP标记来自P0-Cre/CAG-CAT-EGFP(P0-EGFP)双转基因小鼠的NCDC,然后分析其在腭黏膜伤口愈合中的作用。作为腭伤口愈合模型,切除P0-EGFP小鼠左侧腭黏膜,在愈合区域检测干细胞标志物和角质形成细胞标志物。从正常腭黏膜提取NCDC,用干细胞培养基预培养14 d,然后分化为角质形成细胞或成骨细胞以分析多能性。伤口愈合过程从第二天的边缘粘膜再生开始,第 28 天整个伤口区域被含有 EGFP 阳性细胞 (NCDC) 的再生粘膜覆盖。EGFP 阳性细胞占愈合口腔粘膜中约 60% 的细胞,其中 65% 表达干细胞标志物 (Sca-1 + 、PDGFR α + ),30% 表达角质形成细胞标志物 (CK13 + )。在培养的腭粘膜细胞测试中,大约 70% 的 EGFP 阳性细胞表达干细胞标志物 (Sca-1 + 、PDGFR α + )。此外,在分化诱导条件下,培养的 EGFP 阳性细胞被成功诱导分化为角质形成细胞和成骨细胞。我们得出结论,NCDC 作为干细胞存在于成人腭组织中,并有可能在伤口愈合过程中分化为各种细胞类型。

GJB2的细胞特异性传递恢复了鼠标中的听觉功能1 DFNB1耳聋的模型,并介导了NHP Cochlea中的适当表达2
由无处不在的启动子驱动的记者。AAV9-PHP.B矢量(AAV-CBA-EGFP)在CBA启动子下表达EGFP 115(绿色),有效地转导了内毛细胞(IHC),外毛细胞(OHCS)116(Magenta)(Magenta),辅助细胞和其他小鼠Cochlea中的细胞。IHC和OHC通过117个荧光腓骨(Magenta)鉴定。 f。用AAV载体转导的细胞在GRES(AAV-GRE-EGFP)的控制下表达EGFP标记基因118。 值得注意的是,当GRE控制119表达时,在毛细胞中未观察到EGFP。 g,h。用AAV载体转导的细胞在调节元件的控制下表达120 mmgjb2.ha(g)或hsgjb2.ha(h)。 比例尺:10μm(E,F),30 121 µm(G,H)。 122IHC和OHC通过117个荧光腓骨(Magenta)鉴定。f。用AAV载体转导的细胞在GRES(AAV-GRE-EGFP)的控制下表达EGFP标记基因118。值得注意的是,当GRE控制119表达时,在毛细胞中未观察到EGFP。g,h。用AAV载体转导的细胞在调节元件的控制下表达120 mmgjb2.ha(g)或hsgjb2.ha(h)。比例尺:10μm(E,F),30 121 µm(G,H)。122


体外转录的机械模型,结合了焦磷酸镁结晶的作用
图5代表延长的绿色荧光蛋白(EGFP),SARS -2 COV -2 SPIKE蛋白受体结合结构域(RBD)和CAS9模型验证的各种组合:IVT长度的多个DNA序列的IVT产量(Rosa等,2022)。关于模型预测的阴影区域是95%的预测间隔(SI第4节)。数据点上的误差线是根据Rosa等人的整个数据集估计的标准偏差的95%置信区间。(2022)。ivt,体外转录。Rosa等人使用的 DNA序列。 是延长的绿色荧光蛋白(EGFP),SARS -CV -2尖峰蛋白受体结合结构域(RBD)和Cas9基因的组合。DNA序列。是延长的绿色荧光蛋白(EGFP),SARS -CV -2尖峰蛋白受体结合结构域(RBD)和Cas9基因的组合。
荧光 Cas9 mRNA 用于富集 CRISPR-...
图 2:使用荧光 Cas9 mRNA 富集基因敲除 A. 对与 mKate2 Cas9 mRNA 和阳性对照 PPIB crRNA:tracrRNA 共电穿孔并根据 mKate2 荧光进行分选的 K-562 细胞群进行错配检测分析。B. 在次优和最优脂质转染条件下,EGFP Cas9 mRNA 分选的 U2OS 细胞群的 FACS 数据。C. 对 EGFP Cas9 mRNA 分选的 U2OS 细胞群进行错配检测分析