XiaoMi-AI文件搜索系统
World File Search Systemelegans

秀丽隐杆线虫
迫切的治疗需求限制了监管机构批准前治疗药物的研究。因此,对批准后的药物进行分析可以提高我们对药物作用机制的理解,并实现更好的治疗。我们通过对线虫秀丽隐杆线虫进行简单的检测,筛选了 1443 种美国食品药品监督管理局 (FDA) 批准的药物库,发现三种化合物会引起形态变化。抗凝剂噻氯匹定和抗真菌剂舍他康唑都会导致药物积聚,导致咽部解剖结构明显扭曲,急性暴露后死亡,而质子泵抑制剂右兰索拉唑会导致蜕皮缺陷,需要在幼虫发育过程中暴露。强大的遗传模型系统中这种易于检测的缺陷主张继续使用各种模型生物探索现有药物,以更好地了解已经为数百万患者开出的药物。© 2020 作者。由 Elsevier Ltd. 出版。这是一篇根据 CC BY-NC-ND 许可协议 ( http://creativecommons.org/licenses/by-nc-nd/4.0/ ) 开放获取的文章。

秀丽隐杆线虫的学习和化学感受
Arrestin 介导的脱敏使秀丽隐杆线虫的神经元内嗅觉辨别成为可能......................................................................................................................................44

秀丽隐杆线虫的转录适应
摘要 转录适应是最近描述的一种现象,其中一个基因的突变会导致相关基因(称为适应基因)的转录调节。在分子水平上,有人提出,突变的 mRNA(而不是蛋白质功能的丧失)激活了这种反应。虽然已经在斑马鱼胚胎和小鼠细胞系中报道了几例转录适应的例子,但尚不清楚这种现象是否在后生动物中都观察到。我们在此报告了秀丽隐杆线虫的转录适应,并发现该过程需要与突变 mRNA 衰变有关的因子,就像在斑马鱼和小鼠中一样。我们进一步发现了对 Argonaute 蛋白和 Dicer 的需求,这些因子与小 RNA 成熟和转运到细胞核中有关。总之,这些结果为秀丽隐杆线虫的转录适应提供了证据,这是一种进一步研究潜在分子机制的有力模型。
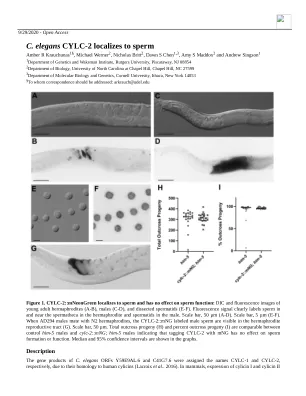
秀丽隐杆线虫 CYLC-2 定位于精子
是睾丸特异性的,免疫组织化学显示蛋白质定位于精子头部的细胞骨架花萼(Hess 等人,1993 年;Hess 等人,1995 年;Rousseaux-Prèvost 等人,2003 年)。为了确定秀丽隐杆线虫 CYLC-1 和 CYLC-2 的定位,我们使用 CRISPR/Cas9 将每个蛋白质内源性地标记为 mNeonGreen (mNG)。我们发现 CYLC-2::mNG 定位于雌雄同体和雄性的精子中(图 1A-F)。检查从雄性解剖的精子细胞显示 CYLC-2::mNG 集中在斑点中(图 1F)。根据它们在精子细胞中的大小和位置,我们预测这些斑点对应于膜状细胞器 (MO)。然而,还需要进一步研究来确认 CYLC-2 是否集中在精子细胞的 MO 中,以及确定精子激活后亚细胞定位是否发生任何变化。

秀丽隐杆线虫用于癌症标志的研究
经过数十年的研究,我们对癌症机制的复杂性的了解(优雅地总结为“癌症的标志”)正在扩大,这种知识带来的治疗机会也在扩大。然而,癌症仍然需要深入研究以减少其巨大影响。在这种情况下,使用简单的模型生物(例如秀丽隐杆线虫),其中发现了凋亡途径的遗传学,可以促进对几种癌症标志的研究。可用于遗传和药物筛查,方便快速有效的基因组编辑,并与伦理动物研究的3RS(“替代,减少和改进”)原理保持一致,秀丽隐杆线虫在揭示癌症机制的复杂网络中起着重要的作用,并在癌症机制中提出了有希望的选择。

18/PTEN 秀丽隐杆线虫突变体
摘要 兴奋和抑制 (E/I) 之间的精细平衡对于大脑正常功能至关重要。GABA 能系统的紊乱会改变这种平衡,是各种神经系统疾病的共同特征,包括自闭症谱系障碍 (ASD)。磷酸酶和张力蛋白同源物 (PTEN) 的突变与 ASD 密切相关,PTEN 是磷脂酰肌醇 3-磷酸激酶/Akt 通路的主要负调节剂。然而,尚不清楚 PTEN 缺陷是否会对抑制和兴奋信号产生不同的影响。利用秀丽隐杆线虫的神经肌肉系统,其中兴奋性 (胆碱能) 和抑制性 (GABA 能) 输入都调节肌肉活动,我们发现 daf- 18 / PTEN 突变会影响 GABA 能(但不影响胆碱能)神经发育和功能。这种选择性影响导致抑制信号传导不足。在 daf- 18/PTEN 突变体中观察到的 GABAergic 系统中的缺陷是由于发育过程中 DAF- 16/FOXO 活性降低所致。生酮饮食 (KGD) 已被证明对与 E/I 失衡相关的疾病有效。然而,其作用机制在很大程度上仍然难以捉摸。我们发现,在早期发育过程中富含酮体 β -羟基丁酸的饮食会诱导 DAF- 16/FOXO 活性,从而改善 daf- 18/PTEN 突变体的 GABAergic 神经发育和功能。我们的研究为 PTEN 突变与神经发育缺陷之间的联系提供了宝贵的见解,并深入探讨了 KGD 潜在治疗效果的潜在机制。

二甲双胍调节骨髓基质细胞在糖尿病小鼠中加速骨骼愈合
抽象映射神经递质身份对神经元是理解神经系统中信息流的关键。它还为研究神经元身份特征的发展和可塑性提供了宝贵的入口点。在秀丽隐杆线虫神经系统中,神经纤维 - 米特的身份在很大程度上是通过编码神经递质生物合成酶或转运蛋白的神经递质途径基因的表达模式分析来分配的。但是,其中许多作业都依赖于可能缺乏相关顺式调节信息的多拷贝记者转基因,因此可能无法提供神经递质使用情况的准确图片。我们分析了秀丽隐杆线虫中所有主要类型的神经递质(谷氨酸,乙酰胆碱,GABA,5-羟色胺,多巴胺,多巴胺,酪胺和章鱼胺)中所有主要类型的神经递质的16个CRIS/CAS9工程敲入报告菌株的表达模式。我们的分析揭示了这些神经递质系统在神经元和神经胶质中以及非神经细胞中的新颖位点,最著名的是在性腺细胞中。所得表达的地图集定义了可能仅是神经肽的神经元,它基本上扩展了能够共同传播多个神经递质的神经元的曲目,并鉴定了单胺能神经植物的新颖位点。此外,我们还观察到单胺能合成途径基因的异常共表达模式,这表明存在新型单胺能发射器。我们的分析导致迄今为止,神经递质使用量最广泛的全动物范围图构成了最广泛的全动物范围图,为更好地理解秀丽隐杆线虫中神经元通信和神经元身份规范铺平了道路。

使用Caenorhabditis elegans
对监管机构负责评估风险的许多化学物质中很少有人对发育神经毒性(DNT)进行了仔细的测试。为加快测试工作以及减少脊椎动物的使用,付出了巨大的努力,致力于替代实验室模型进行测试。DNT的主要机制是由于神经发育过程中化学暴露而改变的神经元结构。Caenorhabditis秀丽隐杆线虫是神经生物学家和发育生物学家广泛研究的线虫,在较小程度上由神经毒理学家进行了研究。秀丽隐杆线虫中神经系统的发育轨迹很容易可视化,通常完全不变并且完全映射。因此,我们假设秀丽隐杆线虫可能是一个强大的体内模型,以测试化学物质,以改变神经元结构的发育模式。为了测试这是否可能是真的,我们开发了一种新型的秀丽隐杆线虫DNT测试范式,其中包括整个发育中的暴露,检查所有主要神经递质神经元类型以进行建筑改变,并测试针对多巴胺能,胆碱能和谷氨酸氨酸性功能的行为。我们使用这种范式来表征早期暴露于发育神经毒性铅,镉和苯并(A)pyrene(BAP)对多巴胺能,胆碱能和谷氨酸氨基氨基氨基氨基甲基体系结构的影响。我们还评估了暴露是否会改变神经元规范,这是通过表达特定神经递质诊断的表达来评估的。我们尚未确定我们检查的神经元明显的神经递质类型发生的情况,但许多神经元形态发生了变化。我们还发现,在秀丽隐杆线虫中,神经元特异性的行为是针对人群中期的秀丽隐杆菌中的,在早期阶段的形态神经退行性变化。功能变化与我们观察到的神经元类型的形态变化一致。我们确定了与哺乳动物DNT文献中报道的变化一致的变化,从而加强了秀丽隐杆线虫作为DNT模型的案例,并进行了新的观察结果,应在以后的研究中进行跟进。

秀丽隐杆线虫中的神经元活动自动分类
秀丽隐杆线虫通常用于研究神经活动,因为 1) 它的基因组和连接组已经得到充分研究,只有 302 个神经元;2) 它的透明度使它能够捕捉详细的神经活动;3) 它具有自体受精的能力,这使得维持基因相同的秀丽隐杆线虫种群成为可能。分析秀丽隐杆线虫的图像具有将特定神经元的活动与行为或环境刺激联系起来的潜力。然而,由于难以命名这些图像中的特定神经元,因此产生此类见解受到限制。当前的方法依赖于手动分类,这既耗时又容易出错。可以利用 ZephIR 等神经元跟踪系统来帮助执行标记。但是,以这种方式使用跟踪系统需要每 11 分钟捕获的图像额外进行 45 分钟的手动标记,才能跟踪特定的神经元。在本研究中,我们开发了一种基于神经网络的分类器,可以自动标记秀丽隐杆线虫中的感觉神经元,准确率高达 91.61%。这是通过使用迭代的、基于地标的神经元识别过程实现的,旨在模仿手动注释程序。

秀丽隐杆线虫 SEL- 5/ AAK1 调节细胞...
摘要 在秀丽隐杆线虫发育过程中,多个细胞会长距离迁移或伸展突起以到达最终位置和/或获得适当形状。Wnt 信号通路是细胞沿前后体轴迁移或细胞生长的主要协调者之一。Wnt 信号的结果受包括内吞作用在内的各种机制的微调。在本研究中,我们发现 SEL-5(哺乳动物 AP2 相关激酶 AAK1 的秀丽隐杆线虫直系同源物)与逆转录复合物一起在 QL 神经母细胞子细胞迁移过程中作为 EGL-20/Wnt 信号的正调节因子发挥作用。同时,SEL-5 与逆转录复合物的协同作用也是排泄道细胞生长所必需的。重要的是,SEL-5 激酶活性不是其在神经元迁移或排泄细胞生长中发挥作用所必需的,并且这两个过程都不依赖于 DPY-23/AP2M1 磷酸化。我们进一步确定,Wnt 蛋白 CWN-1 和 CWN-2 与 Frizzled 受体 CFZ-2 一起正向调节排泄细胞生长,而 LIN-44/Wnt 和 LIN-17/Frizzled 一起产生抑制其延伸的停止信号。