XiaoMi-AI文件搜索系统
World File Search Systemetal
Alval Ulegs蚂蚁地雷。
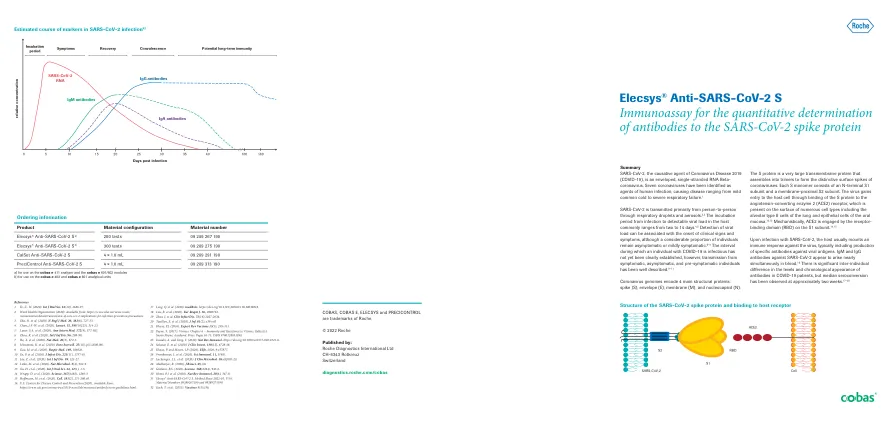
SARS-COV-2尖峰蛋白的结构,并与宿主受体参考文献结合1 Ye,Z.-W。 (2020)。 Int J Biol Sci。 16(10),1686-97。 2个单词健康组织(2020)。 可从:https://www.who.int/news-room/注释/详细信息/详细信息/toxpains/toxpription/toxplion-of-sars-cov-2-implications-for-infote-infection-infection-prevention-prevention-prevention-precapainestions。 3 Zhu,N。等。 (2020)。 n Engl J Med。 20,382(8),727-33。 4 Chan,J.F.-W。等。 (2020)。 柳叶刀。 15,395(10223),514-23。 5 Lauer,S.A。等。 (2020)。 Ann Intern Med。 172(9),577-582。 6 Zhou,R。Etal。 (2020)。 int j inf。 96,288-90。 7 He,X。等。 (2020)。 nat Med。 26(5),672-5。 8 Mizumoto,K。等。 (2020)。 欧元监视。 25(10),PII = 2000180。 9 Gao,M。等。 (2020)。 respir med。 169,106026。 10 Yu,P。等。 (2020)。 J感染。 221(11),1757-61。 11 Liu,Z。等。 (2020)。 int j inf。 99,325-27。 12 Letko,M。等。 (2020)。 NAT微生物。 5(4),562-9。 13 Xu,H。等。 (2020)。 int J Oral Sci。 24,12(1),1-5。 14 Crapp,D。等。 (2020)。 科学。 367(6483),1260-3。 15 Hoffmann,M。等。 (2020)。 单元格。 181(2),271-280.e8。SARS-COV-2尖峰蛋白的结构,并与宿主受体参考文献结合1 Ye,Z.-W。 (2020)。Int J Biol Sci。16(10),1686-97。2个单词健康组织(2020)。可从:https://www.who.int/news-room/注释/详细信息/详细信息/toxpains/toxpription/toxplion-of-sars-cov-2-implications-for-infote-infection-infection-prevention-prevention-prevention-precapainestions。3 Zhu,N。等。 (2020)。 n Engl J Med。 20,382(8),727-33。 4 Chan,J.F.-W。等。 (2020)。 柳叶刀。 15,395(10223),514-23。 5 Lauer,S.A。等。 (2020)。 Ann Intern Med。 172(9),577-582。 6 Zhou,R。Etal。 (2020)。 int j inf。 96,288-90。 7 He,X。等。 (2020)。 nat Med。 26(5),672-5。 8 Mizumoto,K。等。 (2020)。 欧元监视。 25(10),PII = 2000180。 9 Gao,M。等。 (2020)。 respir med。 169,106026。 10 Yu,P。等。 (2020)。 J感染。 221(11),1757-61。 11 Liu,Z。等。 (2020)。 int j inf。 99,325-27。 12 Letko,M。等。 (2020)。 NAT微生物。 5(4),562-9。 13 Xu,H。等。 (2020)。 int J Oral Sci。 24,12(1),1-5。 14 Crapp,D。等。 (2020)。 科学。 367(6483),1260-3。 15 Hoffmann,M。等。 (2020)。 单元格。 181(2),271-280.e8。3 Zhu,N。等。(2020)。n Engl J Med。20,382(8),727-33。4 Chan,J.F.-W。等。 (2020)。 柳叶刀。 15,395(10223),514-23。 5 Lauer,S.A。等。 (2020)。 Ann Intern Med。 172(9),577-582。 6 Zhou,R。Etal。 (2020)。 int j inf。 96,288-90。 7 He,X。等。 (2020)。 nat Med。 26(5),672-5。 8 Mizumoto,K。等。 (2020)。 欧元监视。 25(10),PII = 2000180。 9 Gao,M。等。 (2020)。 respir med。 169,106026。 10 Yu,P。等。 (2020)。 J感染。 221(11),1757-61。 11 Liu,Z。等。 (2020)。 int j inf。 99,325-27。 12 Letko,M。等。 (2020)。 NAT微生物。 5(4),562-9。 13 Xu,H。等。 (2020)。 int J Oral Sci。 24,12(1),1-5。 14 Crapp,D。等。 (2020)。 科学。 367(6483),1260-3。 15 Hoffmann,M。等。 (2020)。 单元格。 181(2),271-280.e8。4 Chan,J.F.-W。等。(2020)。柳叶刀。15,395(10223),514-23。5 Lauer,S.A。等。 (2020)。 Ann Intern Med。 172(9),577-582。 6 Zhou,R。Etal。 (2020)。 int j inf。 96,288-90。 7 He,X。等。 (2020)。 nat Med。 26(5),672-5。 8 Mizumoto,K。等。 (2020)。 欧元监视。 25(10),PII = 2000180。 9 Gao,M。等。 (2020)。 respir med。 169,106026。 10 Yu,P。等。 (2020)。 J感染。 221(11),1757-61。 11 Liu,Z。等。 (2020)。 int j inf。 99,325-27。 12 Letko,M。等。 (2020)。 NAT微生物。 5(4),562-9。 13 Xu,H。等。 (2020)。 int J Oral Sci。 24,12(1),1-5。 14 Crapp,D。等。 (2020)。 科学。 367(6483),1260-3。 15 Hoffmann,M。等。 (2020)。 单元格。 181(2),271-280.e8。5 Lauer,S.A。等。(2020)。Ann Intern Med。 172(9),577-582。 6 Zhou,R。Etal。 (2020)。 int j inf。 96,288-90。 7 He,X。等。 (2020)。 nat Med。 26(5),672-5。 8 Mizumoto,K。等。 (2020)。 欧元监视。 25(10),PII = 2000180。 9 Gao,M。等。 (2020)。 respir med。 169,106026。 10 Yu,P。等。 (2020)。 J感染。 221(11),1757-61。 11 Liu,Z。等。 (2020)。 int j inf。 99,325-27。 12 Letko,M。等。 (2020)。 NAT微生物。 5(4),562-9。 13 Xu,H。等。 (2020)。 int J Oral Sci。 24,12(1),1-5。 14 Crapp,D。等。 (2020)。 科学。 367(6483),1260-3。 15 Hoffmann,M。等。 (2020)。 单元格。 181(2),271-280.e8。Ann Intern Med。172(9),577-582。6 Zhou,R。Etal。 (2020)。 int j inf。 96,288-90。 7 He,X。等。 (2020)。 nat Med。 26(5),672-5。 8 Mizumoto,K。等。 (2020)。 欧元监视。 25(10),PII = 2000180。 9 Gao,M。等。 (2020)。 respir med。 169,106026。 10 Yu,P。等。 (2020)。 J感染。 221(11),1757-61。 11 Liu,Z。等。 (2020)。 int j inf。 99,325-27。 12 Letko,M。等。 (2020)。 NAT微生物。 5(4),562-9。 13 Xu,H。等。 (2020)。 int J Oral Sci。 24,12(1),1-5。 14 Crapp,D。等。 (2020)。 科学。 367(6483),1260-3。 15 Hoffmann,M。等。 (2020)。 单元格。 181(2),271-280.e8。6 Zhou,R。Etal。(2020)。int j inf。96,288-90。7 He,X。等。 (2020)。 nat Med。 26(5),672-5。 8 Mizumoto,K。等。 (2020)。 欧元监视。 25(10),PII = 2000180。 9 Gao,M。等。 (2020)。 respir med。 169,106026。 10 Yu,P。等。 (2020)。 J感染。 221(11),1757-61。 11 Liu,Z。等。 (2020)。 int j inf。 99,325-27。 12 Letko,M。等。 (2020)。 NAT微生物。 5(4),562-9。 13 Xu,H。等。 (2020)。 int J Oral Sci。 24,12(1),1-5。 14 Crapp,D。等。 (2020)。 科学。 367(6483),1260-3。 15 Hoffmann,M。等。 (2020)。 单元格。 181(2),271-280.e8。7 He,X。等。(2020)。nat Med。26(5),672-5。8 Mizumoto,K。等。(2020)。欧元监视。25(10),PII = 2000180。 9 Gao,M。等。 (2020)。 respir med。 169,106026。 10 Yu,P。等。 (2020)。 J感染。 221(11),1757-61。 11 Liu,Z。等。 (2020)。 int j inf。 99,325-27。 12 Letko,M。等。 (2020)。 NAT微生物。 5(4),562-9。 13 Xu,H。等。 (2020)。 int J Oral Sci。 24,12(1),1-5。 14 Crapp,D。等。 (2020)。 科学。 367(6483),1260-3。 15 Hoffmann,M。等。 (2020)。 单元格。 181(2),271-280.e8。25(10),PII = 2000180。9 Gao,M。等。(2020)。respir med。169,106026。10 Yu,P。等。(2020)。J感染。221(11),1757-61。11 Liu,Z。等。 (2020)。 int j inf。 99,325-27。 12 Letko,M。等。 (2020)。 NAT微生物。 5(4),562-9。 13 Xu,H。等。 (2020)。 int J Oral Sci。 24,12(1),1-5。 14 Crapp,D。等。 (2020)。 科学。 367(6483),1260-3。 15 Hoffmann,M。等。 (2020)。 单元格。 181(2),271-280.e8。11 Liu,Z。等。(2020)。int j inf。99,325-27。12 Letko,M。等。 (2020)。 NAT微生物。 5(4),562-9。 13 Xu,H。等。 (2020)。 int J Oral Sci。 24,12(1),1-5。 14 Crapp,D。等。 (2020)。 科学。 367(6483),1260-3。 15 Hoffmann,M。等。 (2020)。 单元格。 181(2),271-280.e8。12 Letko,M。等。(2020)。NAT微生物。5(4),562-9。13 Xu,H。等。(2020)。int J Oral Sci。24,12(1),1-5。14 Crapp,D。等。(2020)。科学。367(6483),1260-3。15 Hoffmann,M。等。(2020)。单元格。181(2),271-280.e8。16美国疾病控制与预防中心(2020年)。 可从:https://www.cdc.gov/coronavirus/2019-ncov/lab/resources/antibody-tests-guidelines.html。16美国疾病控制与预防中心(2020年)。可从:https://www.cdc.gov/coronavirus/2019-ncov/lab/resources/antibody-tests-guidelines.html。

可持续发展报告 - Dell
正如我们上一份报告所指出的,我们认识到,虽然环境管理是企业的重要职责,但我们的责任还延伸到本报告读者感兴趣的其他领域。我们已选择将报告从年度环境报告演变为年度可持续发展报告,这表明我们意识到我们在确保公司影响力和长期生存能力方面发挥着关键作用。我们还改变了报告周期,将本报告的发布与我们的年度回顾发布保持一致,年度回顾可在 http://www.dell.com 上在线获取。我们了解,我们的财务业绩以及我们的环境和社会影响是“三重底线”报告中高度相关的部分。虽然我们并不声称已经实现了此类报告的目标,但我们相信,我们在将这些关键指标整合并纳入报告方法方面取得了真正的进展。

银屑病关节炎治疗的最新进展
简介 银屑病关节炎 (PsA) 的特征是肌腱和滑膜关节炎症(中轴和/或周围型),与皮肤或指甲银屑病的现患、个人或家族病史和/或肌骨外表现 (EMM) 相关,包括指炎、炎症性肠病 (IBD) 和葡萄膜炎。1 它属于脊柱关节病 (SpA) 大家族的一部分,与中轴脊柱关节炎 (axSpA)、肠病关节炎和反应性关节炎一起,所有这些疾病都有重叠的临床、生化和遗传特征。2 近年来,几种新的生物制剂和靶向合成的抗风湿药物 (b/tsDMARD) 已获批用于治疗 PsA3;然而,这些药物并非普遍有效,并且可能与致残性副作用有关。除此之外,全球人口老龄化和多重疾病的增加可能会对 PsA 产生特别深远的影响,因为 PsA 本身是心脏代谢和心理社会疾病的风险因素,而所有这些疾病反过来又会影响

软件公司如何处理人工智能伦理问题?差距分析
公众和学术界对人工智能 (AI) 伦理的讨论正在加速,公众对人工智能伦理问题(例如这些系统中的数据隐私)的认识也越来越高。为了指导人工智能系统的伦理发展,政府和机构参与者以及公司起草了各种人工智能伦理指南。尽管这些指南越来越普遍,但它们因对工业实践缺乏影响力而受到批评。该领域的研究和实践之间似乎存在差距,但其确切性质仍不得而知。在本文中,我们通过将 39 家使用人工智能系统的公司的实践与“可信赖人工智能伦理指南”中提出的可信赖人工智能的七个关键要求进行比较,对当前的技术水平进行了差距分析。本文的主要发现是人工智能伦理指南与实践之间确实存在显著差距。尤其是考虑到软件开发的新要求、社会和环境福祉以及多样性、非歧视和公平性的要求,公司并没有加以解决。

PJM领土中的互连成本分析
在2021年底,PJM有259吉瓦(GW)的发电和存储能力积极寻求网格互连。PJM队列中的容量以太阳能(116 GW)为主导,并且在较小程度上是独立的电池存储(42 GW),太阳能电池混合动力车(32 GW)和Wind(39 GW)。pjm的队列还包含不再寻求互连的项目的数据,包括在使用(79 GW)和已撤回申请的项目(432 GW)(Rand etal。2022)。PJM的队列近年来激增,与2019年年底相比,2021年的活跃队列增加了240%。与互连请求相关的容量几乎是近年来PJM的峰值负载(约155 GW)的两倍,如果建立了大量份额,它可能会对现有发电施加竞争压力。但从历史上看,大多数项目撤回:只有27%的项目要求从2000年到2016年进行互连,从而在2021年期前实现了商业运营。自2012年以来,PJM已实施了许多改革,以减少延迟和项目的取消,包括队列群集扩展(避免队列研究重叠和相关的还算),并为20兆瓦以下的项目(Caspary etal。div>2021)。在2021年,随着互连请求的大幅度增加和多个互连过程研讨会,PJM开始了FERC最近批准的队列改革(FERC 2022)。核心变化的目标是更快,更有效的互连过程,并具有更大的成本确定性。它们包括一种集群,“先准备的,第一服务”方法,基于尺寸的研究存款以及增加的准备存款,这些矿床在研究过程中稍后退出时处于危险之中。为了清除现有的请求积压,PJM将在过渡期间采用“加急流程”,允许在快速轨道上研究其网络升级低于500万美元的项目。展望未来,不促进网络升级需求的项目将能够更快地遵守“加速程序”(PJM 2022B)的最终互连协议。PJM还在2022年推出了一种新的公共工具(队列隔板),以促进在提交互连请求之前对拟议发电的电网影响的评估,但信息仅限于线路加载更改,不包括潜在的升级成本(PJM 2022E)。

构建法国野生和本地来源的本土植物材料供应链:对生态修复和基于自然的解决方案的贡献
欧洲政策旨在同时应对生物多样性和气候危机,部分是通过实施基于自然的解决方案 (NbS)。与此同时,2021-2030 年联合国生态系统恢复十年和越来越多的科学界呼吁在生态恢复计划中优先考虑本地物种。特别是,使用野生和当地来源的本地植物材料对农业生态学、生态恢复和城市环境管理产生了重要益处,并满足了 IUCN NbS 的五项基本标准。随着欧洲越来越多地发起生产和使用野生和当地来源的本地植物材料的倡议,2015 年在法国创建了“V 'eg 'etal local”商标。作为其启动的一部分,11 个生物区被设计为通过控制整个供应链(即从采集到贸易)中植物材料的可追溯性来保证野生和当地来源。此类植物材料已被纳入 NbS 相关计划的主流,本文介绍了其如何帮助实现一系列与 NbS 相关的环境和社会成果。项目经理应继续依靠野生和本地来源的本地植物材料来开展进一步的 NbS 计划并实现其目标。

航天器信息学-ODU数字共享
航天器信息学是近年来最令人兴奋和当代的研究主题之一。许多国家 /地区都在深空探索中部署相关技术,例如AI,机器人技术,机器学习等。此外,需要考虑在航天器,高级技术中涉及的高复杂性,高成本和高风险,需要采用信息建模,模拟,优化和决策支持方法,以提高空间操作的有效性,效率,可靠性和安全性(Du et etal。div>>2017; Rui等。2014)。新兴的信息学方法为航天器领域提供了有关轨道内的航天器,卫星,任何类型的空间探索任务的空间站,从地面控制,用户有效载荷,遥感和条件,遥感和远程感应以及更多的空间自发和计划,预测,预测,计划和控制的活动和活动活动。为了贡献当前和未来的太空探索和航天器的开发,在本期特刊中,我们收集了航天器信息学研究的出色研究论文。每篇论文都经过了匿名专家裁判的Inde Pendent的双盲同行评审。在审查过程之后,接受了八篇高质量论文,并在本期中发表。

使用卷积神经网络的多跳图注意模块(MGA)预测
样本),2)验证数据集(419个样本),3)测试数据集(418个样本)。我们首先检查了MGA的关键参数,即Hop尺寸M,斑块比γ,分支K的数量和多跳权重量β,其中结果显示在图2中。图2(a)表明,M <5的MGA测试MAS低于多头自我注意力(MSA)的MAE,这表明仅考虑重要的嵌入而不是全部计算自我注意事项系数时,这是有益的。最终网络是根据验证数据集的性能选择的。我们还将我们的模型与5种不同的CNN模型进行了比较(Peng etal。,2021)和tsan(Cheng等人,2021)是大脑年龄预测场中的状态模型。在图3(1)中,MGA-SSE-RESNET18在比较中获得了最低的MAE(2.822年)和最高的PCC(0.968)。还评估了其他预测模型,例如视觉变压器(VIT)或图形注意网络(GAT),但表现较差,大概是由于训练数据不足所致。还表明,实施MGA模块会减少模型偏差和方差(图3(2))。从结果中,我们表明,将MGA与常规CNN交织可以提高准确性,从而有效地对脑年龄预测有效。

人工智能代理的可见性
越来越多地将商业、科学、政府和个人活动委托给人工智能代理(能够在有限监督下实现复杂目标的系统),可能会加剧现有的社会风险并引入新的风险。理解和减轻这些风险涉及严格评估现有的治理结构、在必要时修改和调整这些结构,以及确保关键利益相关者的责任。有关某些人工智能代理在何处、为何、如何以及由谁使用的信息(我们称之为可见性)对于这些目标至关重要。在本文中,我们评估了三类提高人工智能代理可见性的措施:代理标识符、实时监控和活动日志记录。对于每一种措施,我们都概述了在侵入性和信息量方面各不相同的潜在实现。我们分析了这些措施如何应用于从集中到分散的部署环境,并考虑到供应链中包括硬件和软件服务提供商在内的各种参与者。最后,我们讨论了我们的措施对隐私和权力集中的影响。进一步了解这些措施并减轻其负面影响有助于为人工智能代理的治理奠定基础。

物种分布建模 - 快速启动指南
限制物种潜在分布的非生物屏障在性质和规模上差异很大。它们可以包括气候,干扰制度和地形属性等因素。大地理量表(例如全球或大陆),据信气候可以控制物种的潜在分布(Araújo和Rozenfeld 2014)。在过去几十年中,已经开发了气候和生物数据的全球数据库,以及广泛的统计和机械物种分布模型(SDMS)。在入侵物种管理中,SDM通常使用全球气候变量,最容易获得的全球环境数据进行参数化,因此有时被称为“气候适应性模型”(Camac等人2024)。这些模型已成为量化潜在占用领域的生物安全性工具,这些工具可以为威胁优先考虑(McGeoch etal。2016),后期监视设计(Camac等人2021,Camac等。2024),以及预期经济影响的估计(Dodd等人2020,Stoeckl等。2023)。BioSecurity Commons为用户提供了广泛的统计SDM功能,该功能允许用户生成地图,以确定环境可能适合基于气候和/或栖息地偏好的物种。对所有SDM的描述也可以在生物安全共享支持门户上找到。