XiaoMi-AI文件搜索系统
World File Search Systemexpress


明确同意基因检测
在您在村庄内窥镜检查中心的治疗过程中,您可能有机会进行遗传测试,如果我们发现似乎是由于特征,大小或数量的特征,大小或数量而可疑的,因此,基于您的年龄,家族病史或其他健康因素,您的医疗保健提供者会感觉到与DNA分析相关的情况,依赖于DNA分析的情况,依赖于癌症,依赖于癌症,依赖于DNA的疾病。提高您的医疗保健提供者对这种测试的医疗益处。基因检测(也称为DNA测试)包括“ DNA分析”,这是指对DNA的医学和生物学检查和分析,以识别体内基因的存在和组成。这种测试着眼于您的染色体,基因和蛋白质。变量可以指示您是否患有某些疾病的风险更高,包括癌症和许多其他疾病。任何此类基因测试的目标是为您的医生和您提供其他信息,以做出有关您的医疗保健的明智选择和决定。因此,基因检测可以是帮助我们提供者提供最佳护理的重要工具。重要的是要记住,所有基因测试都是可选的,除非法律另有规定要求,否则将不会向您,您的医师或授权医疗保健提供者以及未经您进一步书面同意的任何其他人以及任何其他个人或实体披露这些结果。您的DNA样品将通过息肉切除术或活检以以下方式收集。该数据包中的信息旨在解释DNA样本的收集,保留,维护和披露的方式或DNA分析结果,以便获得您的明确同意。我们的提供者可以回答您对基因测试可能遇到的任何问题,以确定是否在本协议下征得您的明确同意。基因检测披露用于基因检测的DNA样品的方式“ DNA样品”定义为可以从中提取DNA或从该样品中提取DNA的任何人类生物学标本。使用和披露DNA样品和DNA分析,将向指定的病理实验室提供DNA样品进行测试,如下所述,标题为“患者明确同意”。将提供给您的医师或授权医疗保健提供者的最终DNA分析,并按照以下指令授权,该指令标题为“患者明确同意”。您的医师或授权的医疗保健提供者将使用DNA分析制定治疗计划。DNA样品和DNA分析的保留和维持DNA样品将由病理实验室保留。DNA分析的结果将由订购医师或授权的医疗保健提供者作为您病历的一部分。



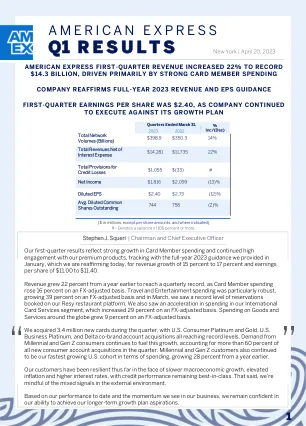
美国运通 - 第一季度业绩
本新闻稿包含《1995 年私人证券诉讼改革法》所定义的前瞻性陈述,这些陈述受风险和不确定性的影响。这些前瞻性陈述涉及美国运通公司目前对业务和财务业绩的预期,包括管理层对 2023 年的展望以及对 2024 年及以后的期望等,其中包含“相信”、“期望”、“预期”、“打算”、“计划”、“目标”、“将”、“可能”、“应该”、“可以”、“会”、“可能”、“继续”等词语和类似表述。请读者不要过分依赖这些前瞻性陈述,这些陈述仅代表陈述作出之日的观点。公司不承担更新或修改任何前瞻性陈述的义务。可能导致实际结果与这些前瞻性陈述存在重大差异的因素包括但不限于以下因素:

表达扫视和视觉注意力
摘要:近年来,眼动研究中最有趣和最有争议的观察之一是猴子和人类的快速扫视现象。这些扫视的反应时间非常短(人类为 100 毫秒,猴子为 70 毫秒),以至于一些眼动专家仍然认为它们是伪像或预期反应,不需要进一步解释。另一方面,一些研究小组认为它们不仅是真实的,而且是研究扫视产生机制、视觉和眼动协调以及视觉注意机制的宝贵手段。这篇目标文章汇集了眼动和相关研究的实验证据 - 特别强调快速扫视 - 以增强我们目前对视觉、视觉注意以及服务于视觉感知和认知的眼动协调的理解。我们假设视运动反射是快速扫视发生的原因,这种快速扫视由与分离的视觉注意和决策有关的高级大脑功能控制。我们提出以神经网络为基础,建立更复杂的数学模型或灵长类动物视运动系统的计算机模拟。

Sarah Glomski - 网站@Duke Express
外科教育与活动实验室 (SEAL),杜克大学 北卡罗来纳州达勒姆 研究生研究员 2024 年 8 月 - 至今 ◦ 设计一个传感器集成组织模型来测量缝合过程中的压力,从而量化外科医生的表现 ◦ 通过创建 ML 算法来对机器人缝合提供实时反馈,从而改善手术机器人的训练 洛斯阿拉莫斯国家实验室 新墨西哥州洛斯阿拉莫斯 / 远程无损检测与评估 (QA) 工程师 2023 年 5 月 - 2024 年 8 月 ◦ 优化核雷管的质量保证,以帮助监测和保存老化的美国核储备 ◦ 开发并实施了一种新的 CT 校准方法,准确度提高了 800%,噪音降低了 500% ◦ 通过创建连接模拟和重建软件的框架来简化 CT 伪影研究 杜克大学伤害生物力学实验室 北卡罗来纳州达勒姆 本科研究员 2021 年 2 月 - 2024 年 5 月 ◦ 训练了几种算法和 ML 模型来对亚脑震荡头部撞击进行分类来自高中足球运动员 ◦ 在年度生物医学工程学会 (BMES) 会议上向数百名工程师介绍了研究结果 Gener8 加利福尼亚州卡尔斯巴德 产品开发实习生 2022 年 6 月 - 2022 年 8 月 ◦ 重启了一个被忽视的采血设备项目,并将血量收集率提高了 400% ◦ 为生物技术客户设计了 60 多个 SolidWorks 零件,并通过幻灯片向利益相关者报告了进展情况 ◦ 获得了产品开发、注塑模具制造、验证测试和管理 BOM 方面的专业知识 DOmath 计算机断层扫描 (CT) 研究,杜克大学 远程本科研究员 2021 年 5 月 - 2021 年 7 月 ◦ 开发并实施了一种新算法,用于更准确地对感兴趣区域进行靶向放射治疗

服务影响-TNT Express
2025年3月12日,对美国的钢铁和铝的进口关税新关税已生效。对原钢和铝进口的关税将为25%,包括半生产产品。一些加工产品也可能受到影响。美国统一关税时间表(HTSU)的特定代码已由商务部发布。请监视更改行业和安全局的网站:https://www.bis.gov/
