XiaoMi-AI文件搜索系统
World File Search Systemgenetics

遗传学
Cas,CRISPR 相关;CRISPR,成簇的规律间隔的短回文重复序列;CRISPRa,CRISPR 介导的转录激活;CRISPRi,CRISPR 介导的转录抑制;crRNA,CRISPR RNA;crRNP,CRISPR 核糖核蛋白;dCas9,核酸酶失活 Cas9;DSB,双链断裂;dsDNA,双链 DNA;dsODN,双链寡脱氧核苷酸;gRNA,向导 RNA;H3K27ac,组蛋白 H3 赖氨酸 27 乙酰化;H3K4me1,组蛋白 H3 赖氨酸 4 单甲基化;LAM-PCR,线性扩增介导的 PCR;LSD1,赖氨酸特异性组蛋白去甲基化酶 1;MCP,MS2 外壳蛋白;MOI,感染复数; p65AD,核因子-κB反式激活亚基激活结构域;PAM,原型间隔区相邻基序;RNAi,RNA干扰;scFV,单链可变片段;sfGFP,超折叠GFP;sgRNA,单向导RNA;ssRNA,单链RNA。



生化遗传学:连接生物化学和遗传学
生化遗传学是一门跨学科领域,融合了生物化学和遗传学,从分子水平上揭示生物过程的复杂性。该领域研究基因如何影响生化过程以及生化途径如何受遗传信息调控。这两门科学学科的融合极大地促进了我们对细胞功能、遗传和疾病机制的理解。历史背景生化遗传学的根源可以追溯到 20 世纪初,当时古典遗传学和代谢途径研究的出现。格雷戈尔·孟德尔等科学家的开创性工作为遗传学奠定了基础,而弗雷德里克·高兰·霍普金斯爵士和亚瑟·哈登等研究人员则探索了酶和辅酶在代谢中的作用。1953 年,詹姆斯·沃森和弗朗西斯·克里克发现 DNA 是遗传物质,这是一个转折点,突出了遗传的分子基础。在随后的几十年里,色谱法、电泳法和分子克隆等技术的发展推动了该领域的发展。这些创新使科学家能够详细分析蛋白质和核酸,从而建立遗传信息和生化功能之间更清晰的联系。

Genetics and beyond
Abstract: Advances in molecular tumor diagnostics have transformed cancer care. However, it remains unclear whether precision oncology has the same impact and transformative nature across all malignancies. We conducted a retrospective analysis of patients with human papillomavirus (HPV)-related gynecologic malignancies who underwent comprehensive molecular profiling and subsequent discussion at the interdisciplinary Molecular Tumor Board (MTB) of the University Hospital, LMU Munich, between 11/2017 and 06/2022. We identified a total cohort of 31 patients diagnosed with cervical (CC), vaginal or vulvar cancer. Twenty-two patients (fraction: 0.71) harbored at least one mutation. Fifteen patients (0.48) had an actionable mutation and fourteen (0.45) received a recommendation for a targeted treatment within the MTB. One CC patient received a biomarker- guided treatment recommended by the MTB and achieved stable disease on the mTOR inhibitor temsirolimus for eight months. Factors leading to non-adherence to MTB recommendations in other patient cases included informed patient refusal, rapid deterioration, stable disease, or use of alternative targeted but biomarker-agnostic treatments such as antibody–drug conjugates or checkpoint inhibitors. Despite a remarkable rate of actionable mutations in HPV-related gynecologic malignancies at our institution, immediate implementation of biomarker-guided targeted treatment recommendations remained low, and access to targeted treatment options after MTB discussion remained a major challenge.
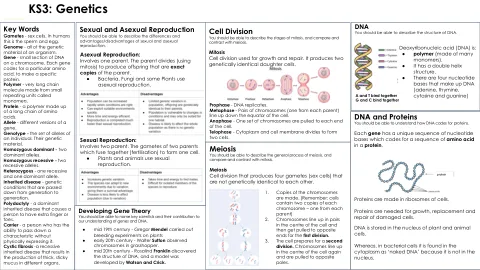
ks3:遗传学
关键词配子 - 性细胞。在人类中,这是精子和鸡蛋。基因组 - 生物体的所有遗传物质。基因 - 染色体上DNA的小部分。每个基因代码特定的氨基酸,以制成特定蛋白质。聚合物 - 由称为单体的小重复单元制成的非常长的链分子。蛋白质 - 由长氨基酸链组成的聚合物。等位基因 - 基因的不同版本。基因型 - 个体的等位基因集。他们的遗传物质。纯合占主导地位 - 两个主要等位基因。纯合隐性 - 两个隐性等位基因。杂合 - 一个隐性和一个主导等位基因。遗传性疾病 - 一代世代相传的遗传疾病。多态度 - 一种主要的遗传疾病,会导致一个人具有额外的手指或脚趾。载体 - 一个能够在不进行物理表达的情况下传递特征的人。囊性纤维化 - 一种隐性遗传性疾病,导致在不同器官中产生厚实的粘性粘液。

PLOS 遗传学
随着物种扩大其地理范围,定居种群面临着新的生态条件,例如新环境和有限的配偶,并因瓶颈和突变负荷积累而遭受人口变化的进化后果。自交通常出现在物种范围边缘,除了抵消配偶的缺乏之外,据推测它还通过增加纯合性和清除来抵抗负荷积累的进化优势。我们研究自交如何通过清除和/或定居速度影响范围扩展过程中遗传负荷的积累。通过模拟,我们解开了由于人口统计学和自交引起的近亲繁殖效应,发现自交者扩张得更快,但仍然会积累负荷,无论交配系统如何。然而,导致这种负荷的变异的严重程度在不同的交配系统中有所不同:较高的自交率会清除大效应的隐性变异,留下较小效应的等位基因负担。我们使用来自避难异交种群和扩大自交种群的全基因组序列,将这些预测与混合交配植物高山南芥 (Arabis alpina) 进行比较。实证结果表明,自交种群中扩张负荷的积累以及清除的证据,与我们的模拟结果一致,这表明虽然清除是自交在范围扩展过程中进化的一项好处,但它不足以防止因范围扩展而导致的负荷积累。


分子遗传学
副教授Mirette Saad是澳大利亚临床实验室的顾问化学病理学家和国家分子遗传学总监。她获得了来自埃及苏伊士运河大学的化学和分子病理学荣誉,以及微生物学亚专业。A/P SAAD从墨尔本大学和Peter Maccallum Cancer Institute获得了NHMRC赞助的癌症遗传学博士学位。,A/P SAAD是AHPRA的注册医生,AHPRA是Australasia皇家病理学家AHPRA(FRCPA),也是澳大利亚临床生物化学家协会(MAACB)的成员。她是RCPA化学病理咨询委员会的主席,RCPA遗传咨询委员会成员,AACB,澳大利亚临床实验室的Precision Medicine Services主席。在临床实验室,A/教授Mirette SAAD领导了非侵入性产前测试(NIPT),产前筛查,个性化药物治疗和癌症的分子基因检测。
