XiaoMi-AI文件搜索系统
World File Search Systemgenomics
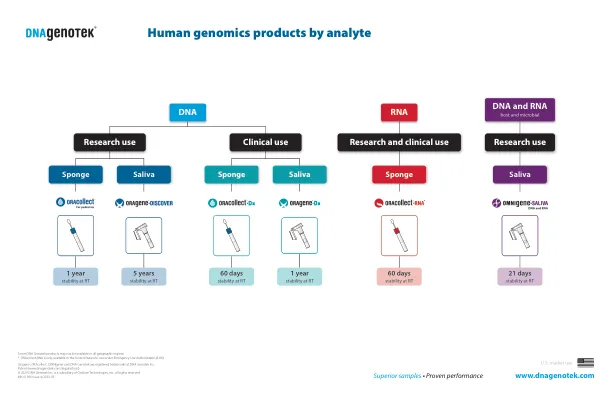
分析物的人类基因组产品
1 Oragene•DX产品手册。 pd-hb-00001 2非侵入性的,辅助收集的高量和质量基因组DNA从幼儿的唾液中。 DNA Genotek。 PD-WP-018 3 Oracollect•RNA产品手册。 DNA Genotek。 pd-hb-00021 4总DNA和RNA产生250μl等分试样的综合剂•根据Oragene•DX人为因素研究的结果,用Qiagen rneasy powermicrobiome kit 5提取的唾液DNA和RNA [K141410和K192920]。1 Oragene•DX产品手册。pd-hb-00001 2非侵入性的,辅助收集的高量和质量基因组DNA从幼儿的唾液中。DNA Genotek。PD-WP-018 3 Oracollect•RNA产品手册。DNA Genotek。pd-hb-00021 4总DNA和RNA产生250μl等分试样的综合剂•根据Oragene•DX人为因素研究的结果,用Qiagen rneasy powermicrobiome kit 5提取的唾液DNA和RNA [K141410和K192920]。

Asteraceae基因组学的综合平台
Asteraceae是最大的被子植物家族,因其出色的药用,园艺和观赏价值引起了广泛的关注。然而,关于星形科植物的研究由于复杂的遗传背景而面临挑战。随着测序技术的持续发展,从星状科物种中积累了大量的基因组和遗传资源。这促使对这个多样化的植物群中对全面的基因组分析的需求。为了满足这种需求,我们开发了Asteraceae基因组学数据库(AGD; http://cbcb.cdutcm.edu.cn/agd/)。AGD充当集中和系统的资源,赋予了各种领域的研究人员,例如基因注释,基因家族分析,进化生物学和遗传育种。AGD不仅包含高质量的基因组序列和细胞器基因组数据,而且还提供了广泛的分析工具,包括BLAST,JBROWSE,SSR FINDER,HMMSEARZER,HMMSEARCH,HEMMAP,HEATMAP,PRIMER3,PLANTIMSISMASH和CRISPRCASFINDER。这些工具使用户能够方便地查询,分析和比较各种星际科中的基因组信息。AGD的建立在推进Asteraceae基因组学,促进遗传育种并通过为研究人员提供全面且用户友好的基因组资源平台来维护生物多样性方面具有巨大的意义。
肿瘤微环境: - 3D Genomics
单细胞多组学技术 • 空间转录组学(10X Visium,通常与 10X Genomics 单核 RNAseq 集成) • 具有大量和少量细胞的单细胞转录组学(分别为 10X Genomics 和 SmartSeq)。 • 来自新鲜冷冻和 FFPE 包埋组织的单核 RNAseq。 • IHC 成像(宽视野/共聚焦、显色/荧光) • 高内涵成像(细胞绘画、HCS) • 单细胞免疫组库分析、B 细胞/T 细胞克隆型研究,通过 FACS 和 MACS 分离细胞类型。 批量细胞技术 • 使用患者来源和对照细胞系在多个治疗领域进行基因表达和药物反应研究的 3D 类器官模型 • 具有多种模态读数的定制细胞检测开发和化合物研究。 • 毛细管印迹 (ProteinSimple) 用于空间转录组学的组织 • 脑、肾、肺、心脏等。 • FFPE 样本即将推出 参考文献
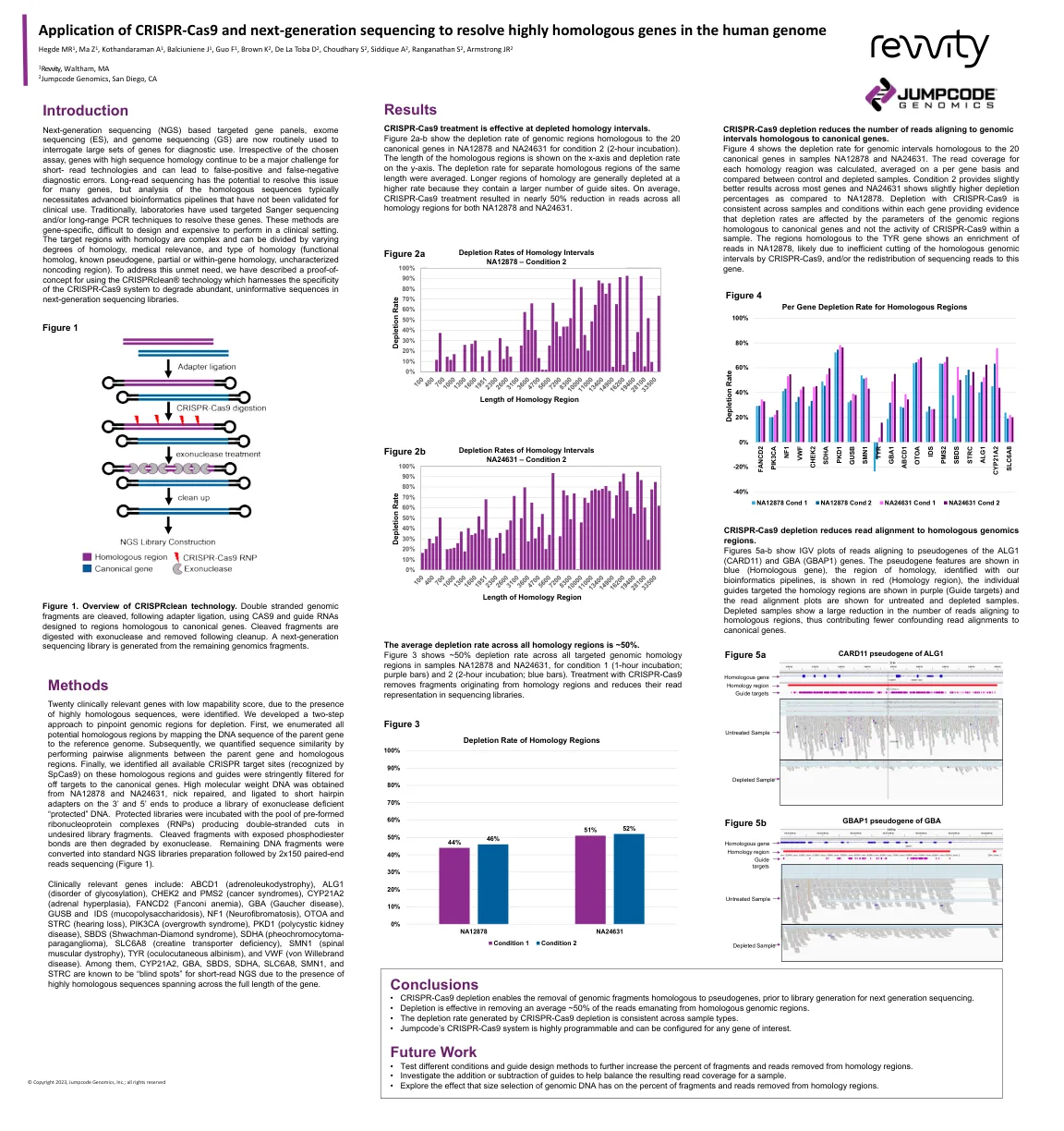
Jumpcode Genomics海报模板
下一代测序(NGS)基于靶向基因面板,外显子组测序(ES)和基因组测序(GS)现在通常用于询问大量基因以供诊断使用。否则对所选的测定法,具有高序列同源性的基因仍然是短读技术的主要挑战,并且可能导致假阳性和假阴性诊断错误。长阅读测序有可能解决许多基因的问题,但是对同源序列的分析通常需要先进的生物信息学管道,这些管道尚未验证用于临床使用。传统上,实验室使用了靶向的Sanger测序和/或远程PCR技术来解决这些基因。这些方法是基因特异性的,难以设计,并且在临床环境中执行昂贵。具有同源性的目标区域是复杂的,可以通过不同程度的同源性,医学相关性和同源性类型(功能同源性,已知的假基因,部分或基因同源性,未表征的非编码区域)进行划分。为了满足这种未满足的需求,我们描述了使用CRISPRCHEAN®技术的概念证明,该技术利用CRISPR-CAS9系统的特异性来降低了下一代测序库中丰富的,无信息的序列。

epizoic seisonidea的基因组学和转录组学(...
背景:Seisonidea(也是Seisonacea或Seisonidae)是一群生活在海洋甲壳动物(Nebalia Spec。)到目前为止仅描述了四个物种。它的单系起源是主要是自由活动的轮动物(单核,bdelloidea)和内寄生虫棘手的蠕虫(acanthocephala)。然而,rotifera-acanthocephala进化枝(rotifera sensu lato或syndermata)内部的系统发育关系受到持续的争论,这是我们对基因组和生活方式如何发展的理解的后果。为了获得新的见解,我们分析了基因组和主要分类单元Seisonidea的转录组的初稿。结果:对GDNA-SEQ和mRNA-SEQ数据的分析发现了法国通道海岸附近的塞森·尼巴里亚·格鲁伯(Seison Nebaliae Grube)的两个遗传学谱系。尽管基因顺序相同,但他们的线粒体单倍型仅具有82%的序列身份。在核基因组中,不同基因紧凑性,GC含量和密码子的用法反映了不同的弦。单倍体核基因组跨越大约。46 MB,其中96%被重建。根据约23,000个超级转录,S。nebaliae中的基因编号应在rotifera-acanthocephala的其他成员发布的范围内。与此相一致,在nebaliae基因组组装中的后唑核直系同源物和ANTP型转录调节基因在所分析的其他组件中相应数量之间。我们还提供了证据表明,旋转 - acanthocephala中seisonidea的基础分支可以反映出对外组的吸引力。因此,通过重建的祖先序列生根,导致了Hemirotifera(bdelloidea+Pararotatoria)内的单系寄生虫(Seisonidea+acanthocephala)。

基因组学在作物改善中的作用
b l meena 1 *,h s meena 1和s p das 2 1 Rapeseed-mustard Research的ICAR导序,拉贾斯坦邦巴拉特布尔,2 iCar-Research for Neh Region Tripura Center,Lembucherra

临床基因组学基础教学大纲
课程名称 临床基因组学基础 学期 2024 年春季 先修和共同必修课程 无 课程负责人 Carolyn Applegate,MGC,CGC 遗传咨询师经理 电子邮件:cdapplegate@jhmi.edu 课程网站 https://jhu.instructure.com/courses/58618 课程描述 本课程提供遗传助理在实验室或诊所工作时使用的遗传学基本背景和工具。主题包括过去和现在遗传对医疗保健的贡献概述、细胞生物学和遗传学、基因检测技术和应用以及临床遗传服务。本课程将以对遗传和基因组学固有的流行文化和道德考虑的讨论结束。本课程旨在概述生物学、医学、基因检测和遗传咨询的各种概念,以便学生掌握背景知识来了解他们特定工作职责。 课程目标 在本课程结束时,您应该能够:

生物医学基因组学分析插件
生物医学基因组学分析插件主要用于癌症和疾病研究,以分析下一代测序 (NGS) 数据。生物医学基因组学分析插件提供各种专业工具、人类和模型物种的参考数据以及全面的模板工作流程集合,涵盖从初始数据处理和质量保证到数据分析、注释和报告的所有步骤。

Genomics 基因体学
■Genomics (基因体学) ■History of DNA sequencing (DNA定序历史) ■The first-generation DNA sequencing (第㇐代DNA定序) ■Next-generation sequencing (NGS,次世代定序) ■CRISPR/cas9 genome editing (基因编辑技术) ■Genome analysis (基因体分析)

Muhammad Sajjad -Lahore
ICAT技术ITRAQ技术医学应用基因组医学;个性化医学生物标志物发现和药物开发中的蛋白质组学。 农业和生物技术中的应用作物改善中的基因组学和蛋白质组学。 生物技术应用:合成生物学,工业酶。 道德,法律和社会影响基因组学和蛋白质组学研究中的道德问题。 数据隐私和同意。 道德,法律和社会影响基因组学和蛋白质组学研究中的道德问题。 数据隐私和同意。ICAT技术ITRAQ技术医学应用基因组医学;个性化医学生物标志物发现和药物开发中的蛋白质组学。农业和生物技术中的应用作物改善中的基因组学和蛋白质组学。生物技术应用:合成生物学,工业酶。道德,法律和社会影响基因组学和蛋白质组学研究中的道德问题。数据隐私和同意。道德,法律和社会影响基因组学和蛋白质组学研究中的道德问题。数据隐私和同意。