XiaoMi-AI文件搜索系统
World File Search SystemhPSC
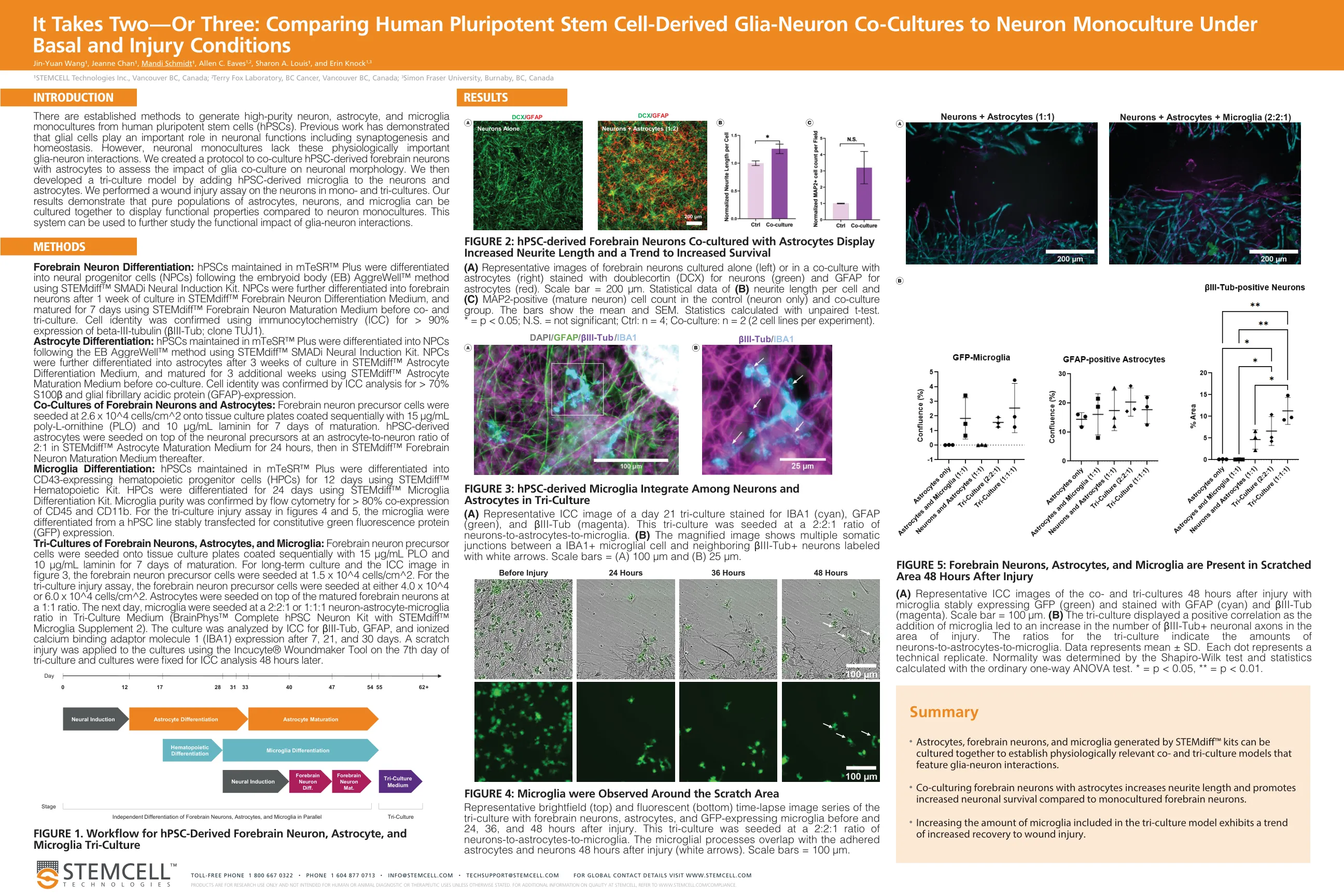
比较人类多能干细胞得出的胶质细胞 -
有建立的方法来产生人类多能干细胞(HPSC)的高纯性神经元,星形胶质细胞和小胶质细胞。先前的工作表明,神经胶质细胞在神经元功能中起重要作用,包括突触发生和稳态。然而,神经元单栽培缺乏这些在生理上重要的神经神经元相互作用。我们创建了一种与星形胶质细胞共同培养HPSC衍生的前脑神经元共同培养的方案,以评估神经胶质共培养对神经元形态的影响。然后,我们通过将HPSC衍生的小胶质细胞添加到神经元和星形胶质细胞中,开发了三个文化模型。我们对单培养的神经元进行了伤口损伤测定法。我们的结果表明,与神经元单栽培相比,可以一起培养星形胶质细胞,神经元和小胶质细胞的纯种群以显示功能特性。该系统可用于进一步研究胶质神经元相互作用的功能影响。

乳酸乳酸乳酸Hkyull 10抑制结直肠肿瘤,并通过其产生的α-甘露糖苷酶恢复肠道菌群人类肠神经系统祖细胞移植改善了赫希斯普伦疾病患者衍生的组织的功能反应
抽象目标赫希斯普伦病(HSCR)是一种严重的先天性疾病,影响1:5000活产。HSCR是由于肠神经系统(ENS)祖细胞在胚胎发育过程中完全定植胃肠道的失败而导致的。这会导致远端肠中炎症病,导致运动活性干扰和蠕动受损。当前,唯一可行的治疗选择是对静脉肠道的手术切除。然而,患者经常患有终身症状,经常需要进行多种外科手术。因此,替代治疗选择至关重要。一种有吸引力的策略涉及人类多能干细胞(HPSC)产生的ENS祖细胞的移植。设计ENS祖细胞是使用加速方案从HPSC生成的,并通过单细胞RNA测序,蛋白质表达分析和钙成像的结合详细介绍了。我们使用器官浴收缩力在体内移植向有机型培养的患者衍生的结肠组织后,测试了ENS祖细胞在HSCR结肠中整合和影响功能反应的能力。结果我们发现,我们的方案始终产生具有早期ENS祖细胞的转录和功能标志的细胞群的高收率。移植后,HPSC衍生的ENS祖细胞在外植的人类HSCR结肠样品中整合,迁移并形成神经元/胶质。与对照组织相比,移植的HSCR组织表现出显着增加的基础收缩活性和对电刺激的反应增加。结论我们的发现首次证明了HPSC衍生的ENS祖细胞在人类HSCR患者结肠组织中重新填充和增加功能反应的潜力。

mTeSR™ Plus
在 mTeSR™ Plus 中培养的 hPSC 的废培养基 pH 值高于在 mTeSR™1 和另一种灵活补料培养基中培养的 hPSC,且细胞密度相似。pH 值和细胞数是在 72 小时不补料后测量的。显示的细胞数范围代表在典型传代过程中观察到的不同密度。这表明在使用 mTeSR™ Plus 进行常规维护期间,可以随时跳过补料两天,同时保持 pH 值高于 7.0。注意:在所有培养基中,在 72 小时不补料之前,培养物的补料量是标准培养基量的两倍,细胞数来自 6 孔板的一个孔。

美国宇航局的高性能太空飞行计算机
HPSC 是一种现代的缓存一致性共享内存多核微处理器,具有八个应用处理核心,使用开放标准 64 位 RISC-V 指令集架构 (ISA) 实现 [5]。HPSC 集成了两个 SiFive X288 核心复合体,每个复合体由 4 个 X280 RISC-V 核心组成。X280 核心采用称为矢量单元的高级功能设计,符合 RISC-V 矢量扩展 (RVV) 标准。矢量单元具有 512 位矢量寄存器长度,支持可变矢量长度计算,最高可达 4096 位宽。RISC-V 矢量是一种强大且超高效的扩展,具有紧凑的代码大小、高性能能力,并且与其他 ISA 青睐的单指令多数据 (SIMD) 架构方法相比,片上 SoC 结构占用的面积有限。此外,RVV 可以在同一软件中使用不同的矢量长度,从而实现可扩展性、灵活性和未来兼容性。

NASA的高性能太空飞行计算机
HPSC是一种现代的高速缓存共享内存多核微处理器,使用开放标准64位RISC-V指令集架构(ISA)[5]实现了八个应用程序处理核心。HPSC集成了两个sifive x288核心复合物,每个复合物由4x x280 RISC-V核组成。X280核心设计的高级功能称为矢量单元,该功能符合RISC-V矢量扩展(RVV)标准。矢量单元具有512位矢量寄存器长度和可变矢量长度计算,最高为4096位。RISC-V向量是一种功能强大且高效的扩展名,具有紧凑的代码大小,高性能功能和ON-DIE SOC结构与单个指令多个数据(SIMD)体系结构方法相比,其他ISA偏爱的soc soc结构的区域有限。此外,RVV可以在同一软件中利用不同的向量长度,从而实现可伸缩性,灵活性和将来的兼容性。

人类胚胎干细胞主要编辑结果的综合分析
Prime editing 是一种多功能且精确的基因组编辑技术,无需供体 DNA 即可将所需的基因修饰直接复制到目标 DNA 位点。该技术在基因功能分析、疾病建模和临床相关细胞(如人类多能干细胞 (hPSC))中的致病突变校正方面具有巨大前景。在这里,我们通过生成一个可由强力霉素诱导的 Prime editing 平台,全面测试了 hPSC 中的 Prime editing。Prime editing 成功诱导了所有类型的核苷酸替换以及小插入和缺失,与其他人类细胞类型中的观察结果相似。此外,我们比较了 Prime editing 和碱基编辑在纠正来自 α 1- 抗胰蛋白酶 (A1AT) 缺乏症患者的诱导多能干细胞中的疾病相关突变方面的表现。最后,全基因组测序表明,与胞嘧啶碱基编辑器的 21 胞苷脱氨酶结构域不同,22 引物编辑器的逆转录酶结构域不会导致基因组中不依赖 RNA 的脱靶突变。23 我们的结果表明,hPSC 中的引物编辑具有补充 24 先前开发的 CRISPR 基因组编辑工具的巨大潜力。25

人类原型多能干细胞通过内源性BMP5/7诱导分化为滋养细胞干细胞,而无需通过幼稚的状态过渡
原代人滋养细胞(TSC)和来自人类多能干细胞(HPSC)的TSC可以在体外对胎盘过程进行模拟。然而,HPSC与TSC的分化涉及的多能状态和因素对TSC的分化知之甚少。In this study, we demonstrate that the primed pluripotent state can generate TSCs by activating pathways such as Epidermal Growth Factor (EGF) and Wingless-related integration site (WNT), and by suppressing tumor growth factor beta (TGFβ), histone deacetylases (HDAC), and Rho-associated protein kinase (ROCK) signaling pathways, all without the addition of exogenous骨形态发生蛋白4(BMP4) - 我们称为TS条件的条件。我们使用时间单细胞RNA测序表征了此过程,以将TS条件与单独使用BMP4激活或与Wnt抑制结合使用的分化方案进行比较。TS条件始终产生一种稳定的增殖细胞类型,该类型紧密模仿了头三年的胎盘细胞增多质细胞,以内源性逆转录病毒基因的激活和缺乏羊膜表达为标志。这是在多个细胞系中观察到的,包括各种引发诱导的多能干细胞(IPSC)和胚胎干细胞(ESC)系。启动衍生的TSC可以在30多个通道中增殖,并进一步指定为多核合胞素粒细胞和跨性滋养细胞细胞。我们的研究表明,在TS条件下,引发HPSC与TSC的分化触发了TMSB4X,BMP5/7,GATA3和TFAP2A的诱导,而无需通过幼稚的

通过局部单倍体人类多能干细胞对 BRCA2 基因变体进行功能注释
BRCA2 基因突变与散发性和家族性癌症有关,可导致基因组不稳定并使癌细胞对聚(ADP-核糖)聚合酶 (PARP) 抑制敏感。本文表明,删除一个 BRCA2 拷贝的人类多能干细胞 (hPSC) 可用于注释此基因的变体并测试其对 PARP 抑制的敏感性。通过使用 Cas9 编辑局部单倍体 hPSC 和从其分化的成纤维细胞中的功能性 BRCA2 等位基因,我们鉴定了该基因中的必需区域以识别允许突变和功能丧失突变。我们还使用 Cas9 直接测试单个氨基酸的功能,包括由意义不明确的临床 BRCA2 变体编码的氨基酸,并鉴定了对用作 BRCA2 缺陷型癌症治疗标准的 PARP 抑制剂敏感的等位基因。局部单倍体人类多能干细胞可以促进基因的详细结构功能分析以及临床观察到的突变的快速功能评估。

使用人多能干细胞衍生的近去中胚层衍生物对膜内骨化的建模
脊椎动物通过两种不同的骨化模式(内膜内和内侧骨术),从三个不同的起源(神经rest,近去中胚层和侧板中胚层)形成其骨骼组织。由于近期中胚层同时会产生膜内和内软骨内骨,因此据认为会引起骨基因生成剂和骨质造基因生成剂。但是,在人类骨骼发育过程中,尚不清楚是什么指导近去中胚层衍生的细胞在不同的骨骼元素中朝着这些不同的命运。要回答这个问题,我们需要实验系统来概括中胚层介导的膜内和内软骨内骨化过程。在这项研究中,我们旨在开发一个基于人类的人体内骨内骨化过程的人类多能干细胞(HPSC)的系统。我们发现,hPSC衍生的近二胚层衍生物的球体培养物会根据刺激产生骨化剂或骨核培养基。前者在小鼠肾胶囊中诱导的膜内骨骼和后者的软骨骨膜。转录pro填充支持以下观点:骨骼特征富含膜内骨状组织。因此,我们开发了一个概括膜内骨术的系统,并通过控制HPSC衍生的副型中胚层衍生物的细胞命运来诱导两种不同的骨化模式。©2023,日本再生医学学会。Elsevier B.V.这是CC BY-NC-ND许可证(http://creativecommons.org/licenses/by-nc-nd/ 4.0/)下的开放访问文章。

Chalcone抑制的心脏中胚层诱导人类多能干细胞的心肌工程
调节性SMAD转录因子(R-SMADS),特别是SMAD 1,5和8。[2]在其磷酸化时,R-SMADS与共同的共肌(SMAD 4)寡聚并转移到核,以调节BMP靶基因的表达。[2b,3] BMP-SMAD信号传导的作用已充分记录在胚胎发生中,尤其是心脏中胚层的形成。[4]在发育中的胚胎中,BMP是从胚外中胚层分泌的,产生形态学的BMP梯度,在浓度,空间和时间下,该梯度指导祖细胞细胞向心脏中胚层的分化。[5]基于胚胎心脏发展的观察结果,在小鼠和人PSC模型中已经开发了采用BMP受体激活的定向分化方案。[4C,6]与这些观察结果一致,我们最近发现,激活蛋白A,BMP4,CHIR99021和FGF2(ABCF-求解)支持心脏中介体形成,包括所有测试的HPSC系(包括胚胎和诱导的Pluripotent semorts),以及在所有测试的HPSC系中,以及随着诱导的PLURIPOTENT的应用 - 心肌。[7]