XiaoMi-AI文件搜索系统
World File Search Systeminducers

提高癌症治疗的精准度:基因治疗和免疫调节在肿瘤学中的作用
基因疗法长期以来一直是治疗罕见病和遗传性疾病的基石,为曾经被认为无法治愈的疾病提供了有针对性的解决方案。随着该领域的发展,其变革潜力现已扩展到肿瘤学,个性化疗法可解决癌症的遗传和免疫相关复杂性。本综述重点介绍了创新的治疗策略,包括基因置换、基因沉默、溶瘤病毒疗法、CAR-T 细胞疗法和 CRISPR-Cas9 基因编辑,重点介绍它们在血液系统恶性肿瘤和实体瘤中的应用。CRISPR-Cas9 是精准医疗的革命性工具,可以精确编辑致癌突变,增强免疫反应并破坏肿瘤生长机制。此外,新兴方法针对铁死亡(一种受调控的铁依赖性细胞死亡形式),为选择性诱导耐药性癌症中的肿瘤细胞死亡提供了新的可能性。尽管取得了重大突破,但肿瘤异质性、免疫逃避和免疫抑制性肿瘤微环境 (TME) 等挑战仍然存在。为了克服这些障碍,人们正在探索新方法,例如双靶向、装甲 CAR-T 细胞以及与免疫检查点抑制剂和铁死亡诱导剂的联合疗法。此外,同种异体“现成” CAR-T 疗法的兴起提供了可扩展且更易于获得的治疗选择。监管环境正在不断发展以适应这些进步,美国的 RMAT(再生医学先进疗法)和欧洲的 ATMP(先进疗法药物)等框架正在快速批准基因疗法。然而,围绕基于 CRISPR 的基因编辑的伦理考虑(例如脱靶效应、种系编辑和确保公平获取)仍然处于最前沿,需要持续的伦理监督。脂质纳米颗粒 (LNP) 和外泌体等非病毒递送系统的进展正在提高基因疗法的安全性和有效性。通过将这些创新与联合疗法相结合并解决监管和道德问题,基因疗法将彻底改变癌症治疗,为血液系统肿瘤和实体肿瘤提供持久、有效和个性化的解决方案。
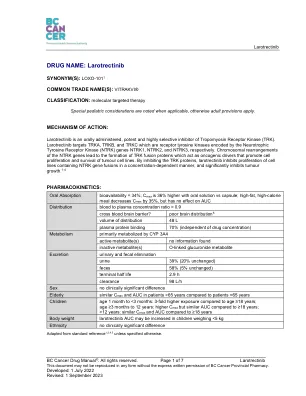
药品名称:Larotrectinib
特别注意事项:注意:•由于神经系统不良事件和疲劳常见,驾驶或操作机器的能力可能会受到影响 2,4 •已有肝功能不全的患者可能需要减少起始剂量 2,4 •与强效 CYP 3A4 抑制剂或诱导剂同时使用时可能需要调整剂量 2,4特殊人群:•与成年患者相比,儿科患者发生 3 级或 4 级中性粒细胞减少症(9-20% vs 0-2%)和体重增加(2-11% vs 0-2%)的发生率更高 2 •65 岁及以上的患者比年轻患者更容易出现疲劳、贫血、头晕、跌倒、步态不稳和低钠血症 2致癌性:未发现信息致突变性:Ames 试验和哺乳动物体外突变试验未发现致突变性。Larotrectinib 在哺乳动物体内染色体试验中不具有致染色体断裂作用。 2,4 生育力:在动物研究中,雌性受试者在暴露量比人类临床暴露量高 8 倍时,黄体减少,发情迟缓发生率增加,子宫重量下降(伴有子宫萎缩)。这些影响是可逆的。在暴露量比人类临床暴露量高 7-10 倍时,未观察到对精子发生或雄性生殖器官组织病理学的影响。2,4 妊娠:在动物研究中,拉罗替尼被证明可以穿过多个物种的胎盘,并在胎儿血液样本中检测到。拉罗替尼在母体毒性剂量以下的暴露量不具有胚胎毒性。然而,在器官形成期间给药时,在母体暴露量比人类临床暴露量高 0.6-9 倍时观察到包括脐膨出和全身水肿在内的畸形。对于有生育能力的女性,建议在开始治疗前进行妊娠测试。对于有生育能力的女性患者和有生育能力的女性伴侣的男性患者,建议在治疗期间以及最后一次服药后至少一个月内采取避孕措施。2,4 由于药物可能会分泌到乳汁中,因此不建议母乳喂养。女性在治疗期间以及最后一次服药后一周内不应进行母乳喂养。2,4

患者:luladur
禁忌症:•用洛拉替尼施用强CYP 3A诱导剂增加了严重的肝毒性的潜力;在启动Lorlatinib之前停止CYP 3A诱导剂(需要一个诱导剂的液latinib(至少等于3个血浆半衰期))。治疗前正确的电解质异常,并监测具有已知危险因素的患者所示的ECG和电解质2•治疗前应充分控制预先存在的高血压2•驾驶或操作机械的能力可能会受到损害,因为神经功能不良事件可能是常见的2致癌性:未进行致癌性研究。2诱变性:在AMES测试中不是诱变。洛拉替尼在体外和体内染色体测试中是哺乳动物的苯二尼。2生育能力:在动物研究中,男性测试受试者的发现包括睾丸/附子症/前列腺,前列腺和睾丸肾小管萎缩的器官重量降低以及附子炎症。效果高2-8倍,部分或完全可逆。治疗前考虑男性患者的生育能力。2,3妊娠:在动物研究中,劳拉替尼引起胚胎毒性。结构畸形(旋转的肢体,畸形的肾脏,圆顶头等。2,3针对育儿潜力的女性患者,建议在治疗前进行妊娠检查。2),胎儿体重的减轻和与人类临床暴露后看到的暴露相似或更少的暴露时观察到植入后丧失。建议在治疗期间和最后一剂劳拉替尼后至少21天进行避孕。洛拉替尼可能与激素避孕药相互作用;因此,建议采用非激素避孕方法。2对于患有怀孕伴侣的男性患者或生育潜力的女性伴侣,在治疗期间和最后一次剂量洛拉替尼后至少3个月,建议采用屏障方法。2,3不建议母乳喂养,因为母乳的潜在分泌。妇女在治疗期间和最后一剂洛拉替尼后不应母乳喂养1周。

靶向遗传毒性和蛋白毒性细胞应激
概要:1。遗传毒性和蛋白质毒性应激在癌变和癌症治疗中的意义…………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………。Proteostasis and proteotoxic stress…………………………………………………………………………………4 1.2.遗传毒性压力……………………………………………………………………………………………………………………………………………在研究遗传毒性和蛋白毒性应激诱导和相关细胞反应的工具中的进步…………………………………………………………………………………………复制应力诱导者是特定遗传毒性应力的来源……………………………………19 2.2。An Advanced Method for QuanƟfying Low-dose DNA Damage and ReplicaƟon Stress Responses……………………………………………………………………………………………………………………………21 2.3.Photo-ManipulaɵOnDNA损伤技术用于细胞研究…………………………………………22 2.4。针对蛋白质毒性应激研究的靶向热蛋白损伤…………………………………………23 2.5。监测毛囊中细胞反应………………………………………………………………二硫代氨基酸盐靶向蛋白质量和DNA修复…………26 3.1。npl4,p97分离酶的适配器,是拆卸纤维的主要目标靶标…………27 3.2。解密的二硫杆的and-canter机制:超越Aldhimhibiɵ…………………………30 3.3。Disulfiram's TargeƟng of NPL4 Impairs DNA ReplicaƟon Dynamics and Induces ATR Pathway MalfuncƟon…………………………………………………………………………………………………………..31 3.4.大麻二醇通过金属硫蛋白途径对二硫杆的效应干扰…………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………3.5。二硫杆重新利用以克服Mulɵple骨髓瘤的抗性………………………………35 3.6。摘要…………………………………………………………………………………………………………………………………………………………癌症疗法中的p97/NPL4途径的新型二硫那甲酸酯络合物造成…………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………3.7。Leveraging Disulfiram, Vorinostat, and PARP inhibitors for CombaƟng CastraƟon-Resistant Prostate cancer……………………………………………………………………………………………………………………38 4.缩写…………………………………………………………………………………………………………………………………………………………………………………………………………………………………………书目……………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………附件1-15………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………… 2…………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………” 4………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………” 6………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………… 8……………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………… 10……………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………” 12……………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………… 14………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………………

癌症干细胞:HCC 领域的潜在突破 - ...
与正常组织中的干细胞一样,癌症干细胞 (CSC) 是肿瘤组织中具有“类干细胞”特征的小细胞群。CSC 具有自我更新和分化为异质性肿瘤细胞的能力,这些肿瘤细胞负责肿瘤的维持和增殖(Batlle and Clevers,2017)。CD34 + /CD138 − 细胞能够在急性髓系白血病中引发肿瘤是 CSC 的第一个确凿证据(Bonnet and Dick,1997)。基于这一突破,随后在多种造血系统癌症和实体瘤中发现了 CSC。肝细胞癌占原发性肝癌发病率的大多数,并且已经通过在 HCC 中鉴定出几种表面标志物证明了 CSC 的存在(Machida,2017)。大量研究表明CSC为HCC提供了增殖、侵袭和复发优势。即便如此,CSC在HCC中的存在仍然存在争议,这在CSC起源理论中尤其明显(见图1)。一些研究表明CSC来源于肝祖细胞(LPC),巨噬细胞分泌的TNF-α在炎症诱导下将LPC转变为CSC为该理论提供了有力证据(LiXF等,2017)。其他研究表明CSC来源于成熟细胞和胆管细胞在遗传和/或表观遗传变化的影响下去分化(Nio等,2017)。更有趣的是,通过多能性诱导物(如 Nanog、Oct4、Yamanaka 因子和 Sox2)重编程产生 CSC 的说法也被广泛接受( Yamashita and Wang,2013 ),也有研究声称 CSC 来源于骨髓干细胞( Kim et al.,2010 )。面对 CSC 来源的争议,研究者尝试利用体外培养和免疫缺陷肿瘤模型探索 CSC 的来源,例如来源于体外培养的球形细胞和来源于癌细胞与干细胞的融合细胞均被认为是 CSC( Wang R. et al.,2016 )。但体外诱导的 CSC 是否与体内肿瘤中的 CSC 一致仍存在疑问( Magee et al.,2012 )。一方面,

束缚反离子导向催化:手性离子对和双功能配体策略在对映选择性金(I)催化中的结合
对映选择性金 (I) 催化的挑战显然与活性配合物的线性几何形状有关,并且在许多情况下与对映决定步骤的外层机制有关。尽管如此,近年来可以通过空间拥挤的配体(其形成嵌入远端活性位点的深手性口袋)、双功能膦或可能通过亲金相互作用形成的双核配合物实现高对映选择性。1 另外,Toste 2 引入了手性反离子策略,其中值得注意的是 BINOL 衍生的磷酸盐在涉及阳离子金中间体的反应中充当手性诱导剂。尽管对于磷酸盐阴离子的确切机制和作用存在一些不确定性,但该策略已显示出突出的潜力,并引发了金 3,4 和其他过渡金属催化的重大进展。 5,6 在金 (I) 催化中,首次公开的分子内氢烷氧基化、氢羧化和氢胺化反应迄今为止仍然是反离子策略的主要应用领域,尽管该方法在理论上应该适用于更广泛的反应。值得注意的是,所有涉及对映体决定步骤中紧密离子对的反应都可能适用,包括那些通过碳阳离子中间体与远程中性金 (I) 单元进行的反应。这种情况可以用图 1.1 中的串联杂环化-亲核加成反应来适当地代表。7 在这种情况下以及其他情况下,手性反离子的立体化学控制受到磷酸盐-碳阳离子对的空间排列不明确和灵活的影响。我们认为可以通过以某种方式将磷酸盐反离子束缚在阳离子金复合物上来克服这个缺点(图 1.2b)。将磷酸单元连接到金配体的共价系链可能为关键中间体提供足够的几何约束和分子组织,从而实现有效的立体化学控制。如果正确实施,这种方法可能会突破对映选择性金催化以及更广泛地说对映选择性过渡金属催化中“离子配对策略”的极限。之前已经报道过在分子内嵌入阴离子的过渡金属配合物。然而在这些

利用分子靶向疗法预防癌症
如今,肿瘤学家就像人体侦探,不再使用放大镜,而是利用分子病理学的新工具,不仅寻找基因或分子靶点,以针对性地开发创新的抗癌疗法,还寻找分子改变,以便识别出有患肿瘤风险的人群,从而进行预防。这就是精准肿瘤学,借助这种精准肿瘤学,如今我们不仅能够基于精准医学模型,通过识别与患癌风险增加相关的特定基因组决定因素,进行治疗,还能进行个性化癌症预防。该领域包括一系列干预措施,以便在早期识别癌症或避免疾病的发生。乳腺癌的 BRCA 突变状态就是一个典型例子:BRCA 基因突变是乳腺癌的风险因素,对于携带该基因突变的女性,可以提供更频繁的乳房筛查计划(作为二级预防的一部分),或者使用芳香化酶抑制剂或抗雌激素治疗来加强一级预防 [ 1 ]。分子病理学是精准肿瘤学的基石,今天,我们有必要学会研究细胞信号转导通路中不仅单个的改变,还要整体的改变。通过这种方式,分子病理学家可以为临床医生提供关键信息,以推动治疗选择。所谓的组织学模型长期以来一直主导着肿瘤学临床研究和临床实践,现在分子模型已成为其重要组成部分 [ 2 – 4 ]。在这种方法中,起点是肿瘤来源的器官,然后进行组织学检查,识别任何分子改变并选择药物,通过选择更有可能对治疗有反应的患者。组织学模型已经被不可知论模型所超越,在不可知论模型中,肿瘤治疗方法是基于特定的基因组改变或特定的分子方面选择的,这些改变或分子方面可能存在于各种肿瘤中,代表细胞靶点。有趣的是,突变模型还包括微生物群,它代表了生活在体内的数十亿微生物的集合,为我们的生命提供基本支持 [ 5 ]。靶向抗癌疗法涉及使用药物通过干扰参与肿瘤进展的分子(称为“分子靶点”)来阻断癌细胞的生长和扩散。靶向抗癌疗法与传统化疗有几个不同之处:(i)靶向抗癌疗法作用于癌细胞的特定分子靶点,而大多数化疗作用于所有快速繁殖的细胞,包括正常细胞和癌细胞;(ii)靶向抗癌疗法阻止肿瘤细胞的增殖(细胞抑制),而化疗药物杀死肿瘤细胞(细胞毒性);(iii)靶向抗癌疗法与其靶点发生特异性相互作用,而许多化疗是根据其细胞毒性能力来确定的。靶向抗癌疗法目前是许多抗癌药物开发的核心,包括激素疗法、信号转导抑制剂、基因表达调节剂、凋亡诱导剂、血管生成抑制剂、免疫疗法和释放毒素的化合物。值得注意的是,靶向癌症疗法在获得性耐药性方面存在一些限制。因此,靶向抗癌疗法联合使用效果最好。例如,最近的一项研究发现,在 BRAF V600E 突变的黑色素瘤中使用两种药物比单独使用一种药物更能减缓耐药性的产生和病情进展[ 6 ]。此外,

质子泵抑制剂与癌症:现状
幽门螺杆菌感染和 Zollinger-Ellison 综合征,以及预防高危患者(年龄 > 65 岁,有胃肠道溃疡病史或同时接受抗血小板、抗凝或皮质类固醇治疗)的非甾体抗炎药 (NSAID) 相关胃肠道病变(Strand 等人,2017 年)。许多此类疾病通常需要长期治疗,这增加了患者发生临床上显著的药物相互作用的可能性。此外,标签外处方已被广泛报道,特别是在功能性消化不良和预防非风险患者 NSAID 引起的胃十二指肠病变方面(Lassalle 等人,2020 年)。自 1980 年代后期推出市场以来,PPI 在许多国家的使用都有所增加。例如,在法国,2015 年有超过 1500 万拥有医疗保险的人(占法国成年人口的近三分之一)是 PPI 使用者(Singh 等人,2018 年;Lassalle 等人,2020 年)。在一项研究中,三分之一的患者无法确定 PPI 指征,四分之三的 NSAID 预防性处方没有发现可测量的风险因素(Lassalle 等人,2020 年)。大约 20% 的癌症患者使用 PPI(Kinoshita 等人,2018 年;Tvingsholm 等人,2018 年;Sharma 等人,2019 年);然而,PPI 通常会被过量用于治疗化疗的副作用(如 GERD)或作为与皮质类固醇或 NSAID 联合治疗的预防措施(Lassalle 等人,2020 年)。总体而言,PPI 被认为不良事件很少,因为它们通常耐受性良好。然而,据报道,PPI 与胃肠道疾病(恶心、腹痛、传输障碍)、离子吸收障碍(低镁血症、铁缺乏、维生素 B12 缺乏)、肾衰竭、感染(肺炎、艰难梭菌感染、腹膜炎)和骨折有关(Singh 等人,2018 年;Yibirin 等人,2021 年)。此外,PPI 还参与各种药物 - 药物相互作用 (DDI) (Wedemeyer 和 Blume,2014 年;Strand 等人,2017 年;Patel 等人,2020 年;Uchiyama 等人,2021 年)。通过提高胃液 pH 值,PPI 会影响胃液 pH 依赖性药物的吸收。事实上,某些弱碱性药物的胃液 pH 值升高会导致溶解度降低,随后的吸收率也会降低 (Wedemeyer 和 Blume,2014 年;Patel 等人,2020 年)。PPI 也可能影响药物消除,因为它们是有机阳离子转运蛋白 (OCT,参与底物药物的肾脏排泄) 和 P-糖蛋白流出转运蛋白的潜在抑制剂 (Wedemeyer 和 Blume,2014 年;Patel 等人,2020 年)。 PPI 主要在肝脏中通过细胞色素 P450 酶 (CYP) 系统代谢,主要是 CYP2C19 和 CYP3A4 ( Wedemeyer and Blume, 2014 )。它们能够作为 CYP 的抑制剂或诱导剂;抑制 CYP 会增加全身对药物的暴露量 (Patel 等人,2020 年)。奥美拉唑对 CYP2C19 具有高亲和力,对 CYP3A4 具有中等亲和力,因此具有相当大的 DDI 潜力 (Wedemeyer and Blume, 2014 年)。埃索美拉唑也能在临床上显著抑制 CYP2C19,而其他 PPI 对 CYP2C19 的抑制在临床上并不重要 (Patel 等人,2020 年)。然而,只有少数涉及 PPI 的 DDI 具有临床意义 (Wedemeyer and Blume, 2014 年)。尽管如此,

了解目标蛋白质降解
1. 什么是蛋白质?它们有什么作用?:MedlinePlus Genetics。访问日期:2021 年 5 月 3 日。https://medlineplus.gov/genetics/understanding/howgeneswork/protein/ 2. Klaips CL、Jayaraj GG、Hartl FU。衰老和疾病中的细胞蛋白质稳态途径。J Cell Biol。2017;217(1):51-63。doi:10.1083/jcb.201709072 3. Ciechanover A。蛋白水解:从溶酶体到泛素和蛋白酶体。Nat Rev Mol Cell Biol。2005;6(1):79-87。doi:10.1038/nrm1552 4. Oprea TI、Bologa CG、Brunak S 等人。人类基因组中未探索的治疗机会。天然药物发现评论。2018;17(5):317-332。doi:10.1038/nrd.2018.14 5. Hopkins AL、Groom CR。可用药基因组。天然药物发现评论。2002;1(9):727-730。doi:10.1038/nrd892 6. Collins I、Wang H、Caldwell JJ、Chopra R。通过调节泛素-蛋白酶体途径进行靶向蛋白质降解的化学方法。Biochem J。2017;474(7):1127-1147。doi:10.1042/BCJ20160762 7. Ito T、Ando H、Suzuki T 等人。确定沙利度胺致畸性的主要靶点。Science。 2010;327(5971):1345-1350。doi:10.1126/science.1177319 8. Krönke J、Udeshi ND、Narla A 等。来那度胺可导致多发性骨髓瘤细胞中 IKZF1 和 IKZF3 选择性降解。Science。2014;343(6168):301-305。doi:10.1126/science.1244851 9. Lu G、Middleton RE、Sun H 等。骨髓瘤药物来那度胺可促进 cereblon 依赖性 Ikaros 蛋白破坏。Science。2014;343(6168):305-309。 doi:10.1126/science.1244917 10. Gandhi AK、Kang J、Havens CG 等。免疫调节剂来那度胺和泊马度胺通过调节 E3 泛素连接酶复合物 CRL4(CRBN.) 诱导 T 细胞阻遏物 Ikaros 和 Aiolos 降解,从而共刺激 T 细胞。Br J Haematol。2014;164(6):811-821。doi:10.1111/bjh.12708 11. Chamberlain PP、Lopez-Girona A、Miller K 等。人类 Cereblon–DDB1–来那度胺复合物的结构揭示了对沙利度胺类似物反应的基础。Nat Struct Mol Biol。2014;21(9):803-809。 doi:10.1038/nsmb.2874 12. Ito T, Handa H. Cereblon 及其下游底物作为免疫调节药物的分子靶点。Int J Hematol。2016;104(3):293-299。doi:10.1007/s12185-016-2073-4 13. Matyskiela ME, Lu G, Ito T 等人。一种新型 cereblon 调节剂将 GSPT1 募集到 CRL4 CRBN 泛素连接酶中。Nature。2016;535(7611):252-257。doi:10.1038/nature18611 14. Chamberlain PP, Cathers BE。Cereblon 调节剂:低分子量蛋白质降解诱导剂。Drug Discov Today Technol。 2019;31:29-34。doi:10.1016/j.ddtec.2019.02.004 15. Baek K、Schulman BA。分子胶概念固化。Nat Chem Biol。2020;16(1):2-3。doi:10.1038/s41589-019-0414-3 16. Scheepstra M、Hekking KFW、van Hijfte L、Folmer RHA。药物发现中用于蛋白质降解的双价配体。Comput Struct Biotechnol J。2019;17:160-176。doi:10.1016/j.csbj.2019.01.006