XiaoMi-AI文件搜索系统
World File Search Systemlog2
BCYC-SITC-2024-BT7480-Mechanistic-Biomarkers.pdf
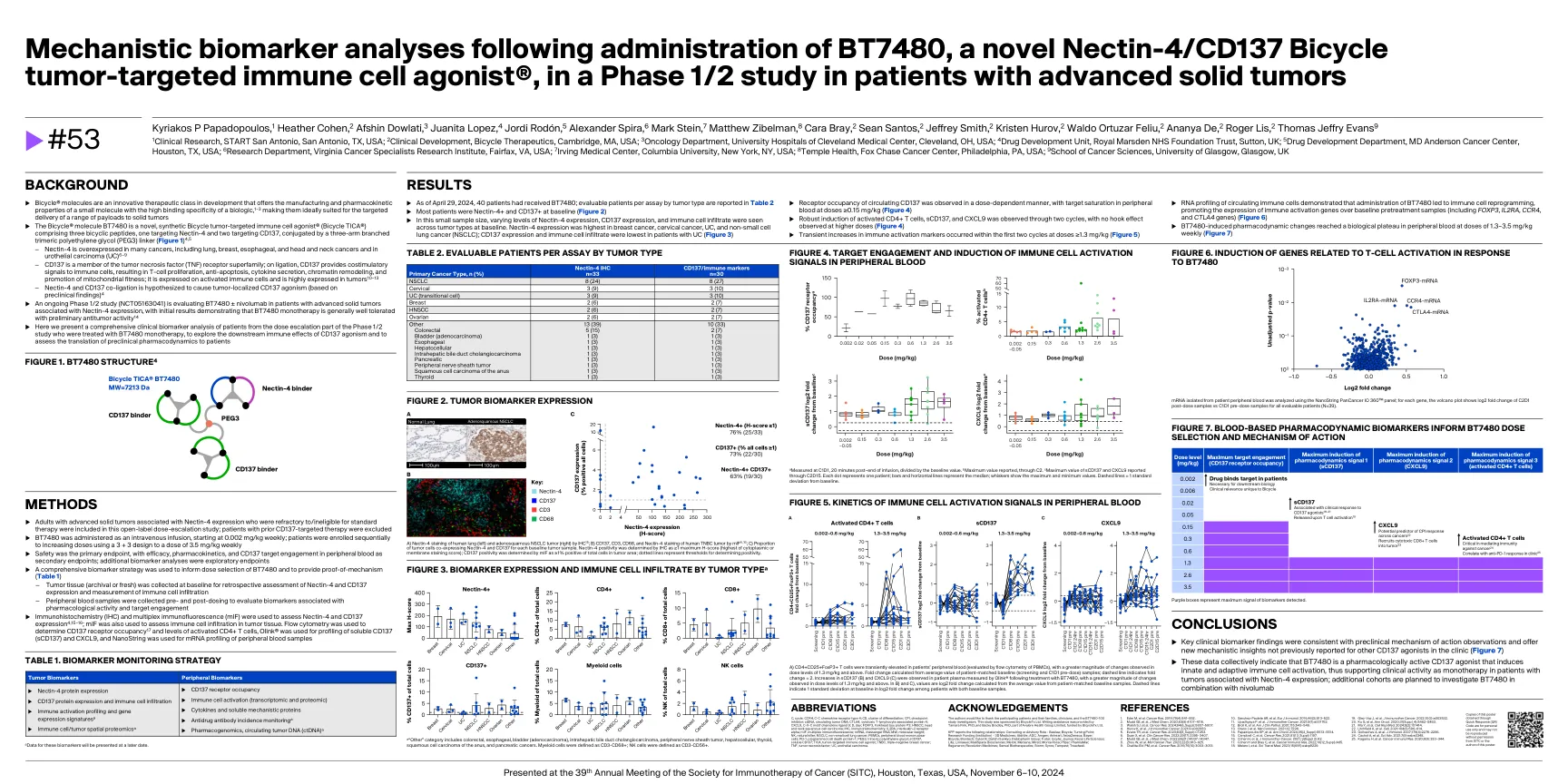
a)患者外周血(通过PBMC的流式细胞仪评估)CD4+CD25+FOXP3+T细胞瞬时升高,并且在1.3 mg/kg及以上的剂量水平上观察到的变化更大。折叠变化是根据患者匹配的基线(筛查和C1D1预剂量)样品的平均值计算得出的;虚线表示折叠变化= 2。在用BT7480治疗后,在通过Olink®测量的患者血浆中观察到SCD137(b)和CXCL9(C)的增加,在剂量水平为1.3 mg/kg及以上时观察到更大的变化。在b)和c)中,是根据患者匹配的基线样本的平均值计算出的log2折叠变化。虚线表示两个基线样品患者的log2折叠变化中基线时的1个标准偏差。

CRISPR-Cas9 筛选用于目标识别。
图 2:PLX-4032 治疗后 sgRNA 计数分布的变化。log2 转换的 sgRNA 计数的箱线图和须线图(基线 - 抗生素选择后的转导细胞;DMSO 和 PLX-4032 - 筛选终点的细胞(治疗 14 天))。箱线图的范围从第一四分位数到第三四分位数,并以黑线显示中位数。此外,下部和上部相邻值显示为须线,异常值显示为圆圈。

高质量亮度分析用于蛋白质和基因表达多重分析
图3。用PHA,CONA或LPS刺激后,基因(RNA)与蛋白质表达的相关性与蛋白质表达的相关性。在刺激后12、24和48小时,ENA(CXCL5),GRO-ALPHA(CXCL1),MCP-3(CCL7)和BLC(CXCL13)的相对RNA和蛋白质表达。将Quantigene plex人免疫反应面板80-plex数据(线图)标准化为管家PPIB。使用Procartaplex人免疫反应面板80-plex获取蛋白质数据。数据(条形图)显示为log2折叠在未刺激的控制样本上的变化。
XMT-2056,一种针对 HER2 的免疫合成刺...
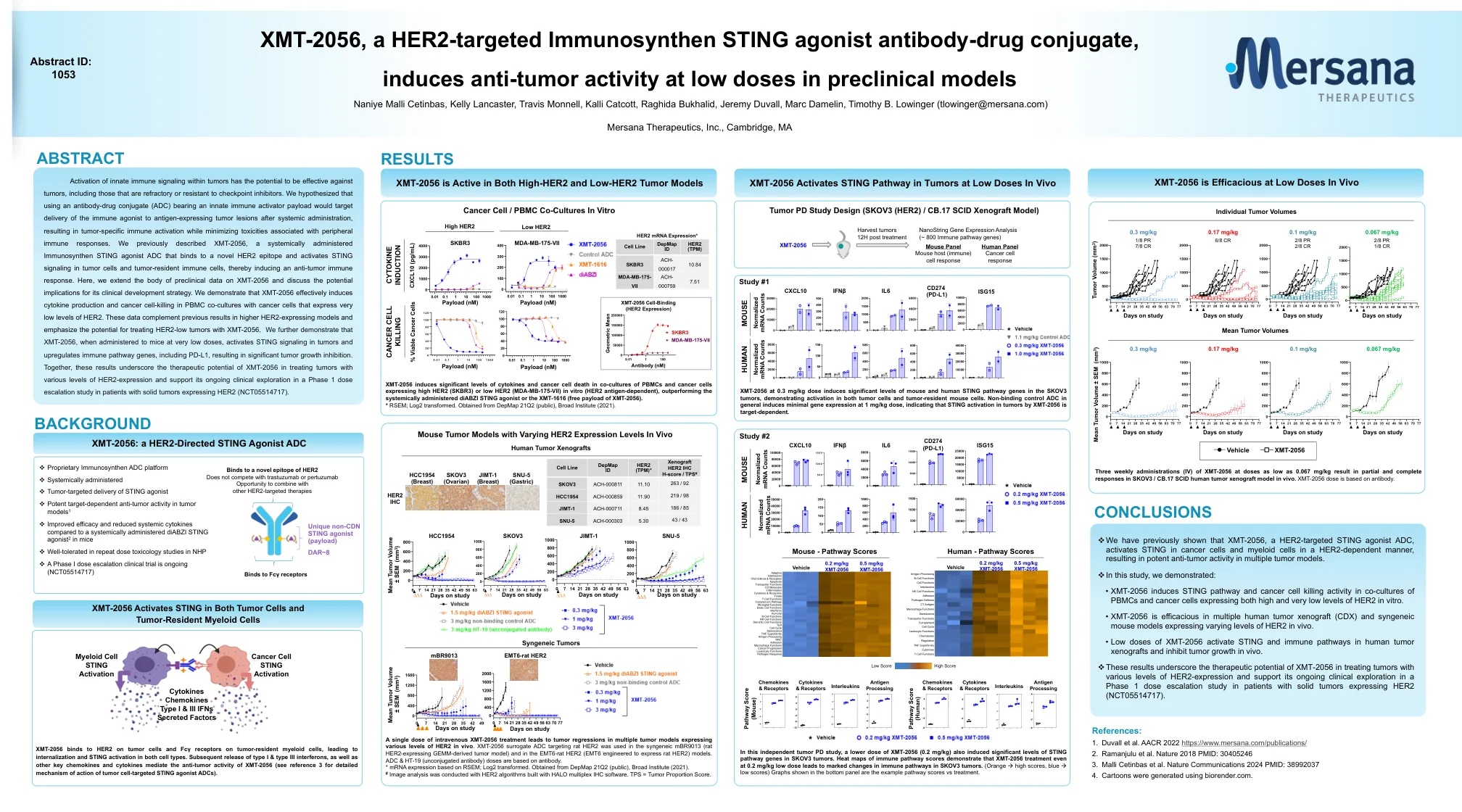
单剂量静脉注射 XMT-2056 治疗可导致体内表达不同水平 HER2 的多种肿瘤模型中的肿瘤消退。靶向大鼠 HER2 的 XMT-2056 替代 ADC 用于同源 mBR9013(表达大鼠 HER2 的 GEMM 衍生肿瘤模型)和 EMT6-大鼠 HER2(经设计表达大鼠 HER2 的 EMT6)模型。ADC 和 HT-19(未偶联抗体)剂量基于抗体。* mRNA 表达基于 RSEM;Log2 转换。来自 DepMap 21Q2(公共),Broad Institute(2021 年)。# 使用 HALO 多重 IHC 软件构建的 HER2 算法进行图像分析。TPS = 肿瘤比例评分。

olink®目标48免疫监视
。预处理过程中的下一步是设置相对于桥接因子桥接不同套件批次之间的数据的值。产生的归一化蛋白质表达(NPX)单位在log2量表上,其中较大的数字代表样品中较高的蛋白质水平,通常为背景水平或接近零。通过使用非线性逻辑回归模型中的四个参数,将NPX值拟合到标准曲线,以描述免疫测定的形状来获得标准浓度单位(PG/mL)中的蛋白质浓度。标准曲线是在验证过程中定义的,并通过产品页面(olink.com/target48human)找到。三个示例如图2所示。

editco-supplied 802-30f IPS单元线
图1。核骨质表明802-30F细胞在基因组上是稳定的。*野生型802-30F细胞(通道19)的Karyostat结果表明基因组完整性已维持。全基因组视图以一个高级副本编号显示了所有体细胞和性染色体。平滑的信号图(右y轴)是log2比的平滑,它描述了微阵列上探针的信号强度。一个值为2表示普通拷贝数状态(CN = 2)。3的值代表染色体增益(CN = 3)。1的值表示染色体损失(CN = 1)。粉红色,绿色和黄色表示每个单独的染色体探针的原始信号,而蓝色信号表示用于识别副本数和畸变的归一化探针信号(如果有)。*改编自Thermo Fisher Scientific的描述。

利用 Qutrit 拓展量子计算机的前沿
中级量子 (NISQ) 计算。NISQ 机制考虑了只有几十到几百个量子比特 (qubits) 和中等误差的近期机器。鉴于量子资源的严重限制,充分优化量子算法的编译对于成功计算至关重要。先前的架构研究已经探索了映射、调度和并行等技术,以扩展可能的有用计算量。在本文中,我们考虑另一种技术:量子三元组 (qutrits)。虽然量子计算通常表示为量子比特的两级二进制抽象,但量子系统的底层物理本质上并不是二进制的。虽然经典计算机在物理层面以二进制状态运行(例如,在阈值电压之上和之下剪切),但量子计算机可以自然访问无限的离散能级谱。事实上,硬件必须主动抑制更高级别的状态才能实现两级量子比特近似。因此,使用三级量子位只不过是选择增加一个离散能级,虽然代价是增加出错几率。先前对量子位(或更一般地,d 级量子位)的研究只发现,扩展量子比特可获得常数因子增益。总体而言,先前的研究 1 强调了量子位的信息压缩优势。例如,N 个量子比特可以表示为 N=log2ð3Þ 量子位,这会导致运行时间有 log2ð3Þ1:6 常数因子改进。我们的方法以一种新颖的方式使用量子位,本质上是使用第三状态作为临时存储,但是代价是每次操作的错误率更高。在这种处理下,运行时间(即电路深度或关键路径)渐近更快,计算的可靠性也得到了提高。此外,我们的方法仅在中间阶段应用量子三元操作:输入和输出仍然是量子位,这对于实际设备上的初始化和测量非常重要。2;3

单细胞转录组学揭示了鼠心力衰竭中成纤维细胞激活的独特模式,并保留的射血分数
a)综合鼠HFPEF和HFREF(ANGII和MI)成纤维细胞研究的示意图。b+c)综合成纤维细胞的UMAP嵌入,疾病(HF,心力衰竭)与对照(B),研究(C)。d)综合成纤维细胞状态的顶部细胞状态标记表达的概述。e)UMAP嵌入,显示由细胞簇着色的集成的成纤维细胞地图集,即集成的成纤维细胞状态(IFS)。标签指示基于功能表征的可能的成纤维细胞分化。f)基于综合成纤维细胞状态中足迹基因的效果大小(AVG log2折叠变化)的估计途径活动。*后代z得分> 2。g)具有综合成纤维细胞状态标记的细胞外基质基因组的过度代表性分析。使用Benjamini Hochberg校正的超几何测试, *Q <0.05,** Q <0.01,*** q <0.001。

Cas9+ 条件性永生化中性粒细胞祖细胞作为全基因组 CRISPR 筛选中性粒细胞分化和功能的工具
图1. 分化后的Cas9+ER-Hoxb8分化中性粒细胞与原代中性粒细胞十分相似。(A)流式细胞术分析分化0、3、4天的Cas9+ER-Hoxb8细胞。(B)在有和没有G-CSF的情况下,分化4天后CD11b和Ly6G染色的流式细胞术分析。(C、D)分化前(C)(D)和分化4天后的Cas9+ER-Hoxb8的TEM。(E)分化2天后Cas9+ER-Hoxb8的基因表达谱与Immunology Genomes数据库中的RNAseq数据进行比较。Y轴表示不同细胞类型的log2(基因表达值/所有基因的平均表达值);从左到右依次为:巨噬细胞(MF_PC、MF_Fem_PC、MF_226+II+480lo_PC、MF_RP_Sp、MF_Alv_Lu、MF_pIC_Alc_Lu、MF_microglia_CNS、MF_AT)、单核细胞(Mo_6C+II-_Bl、Mo_6C-II-)、中性粒细胞(GN_BM、GN_Sp、GN_Thio_PC)和肥大细胞(MC_heparainase_PC)。所有数据代表至少 2 次实验。

胚胎横纹肌肉瘤中的 ERBB2(HER2)扩增:罕见病例中的潜在治疗目标?
图 1 两例 ERBB2 扩增的横纹肌肉瘤 (RMS) 的形态学、免疫组织化学 (IHC) 和遗传特征。 (A) 病例 1 中 ERBB2 扩增子范围的全基因组视图 (顶部) 和详细视图 (底部)。 (B) Circos 图描绘了 17 号染色体 (病例 1) 中的结构变异。请注意 17q 染色体臂中两个扩增子之间的交换。17q 中的两个扩增子以红色注释。 (C) IHC 显示病例 1 (左) 和病例 2 (右) 中 HER2 (ERBB2) 蛋白的细胞质表达强烈。 (D) 17 例儿童 RMS 中 ERBB2 的 mRNA 表达水平;两例 ERBB2 扩增的病例的表达值比无扩增的 RMS 高 50 倍以上。y 轴显示 log2 转换中的表达值。 (E)对病例 1 的培养细胞的间期细胞核进行荧光原位杂交 (FISH),表明扩增的序列被组织成双微体 (dmin)