XiaoMi-AI文件搜索系统
World File Search Systemlz
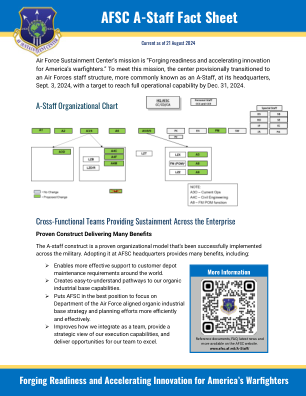
AFSC A 员工情况说明书 - 空军支援中心
人事局 ( DP ) 已改为 A1 ,后勤局 ( LG ) 连同其原有部门一起被拆分,办公室被移至相应的 A3/4 和 A5/8/9 局,情报局 ( IN ) 现为 A2 。 工程局 ( EN )、财务管理局 ( FM )、承包局 ( PK ) 和软件局 ( SW ) 保留,但不采用 A 员工编号。 人事执行局 ( DZ )、财务管理执行局 ( FZ ) 和后勤执行局 ( LZ ) 已撤销。 飞机维护 ( AS )、参谋长 ( DS )、历史学家 ( HO )、监察长 ( IG )、信息保护 ( IP )、军法署 ( JA )、公共事务 ( PA )、小企业 ( SB ) 和安全 ( SE ) 等较小的办公室都属于新的特殊职员类别。

基于反射型超表面的红外卡塞格林望远镜设计与数值分析
1 中国科学院微电子研究所微电子仪器与设备研发中心,北京市北土城西路 3 号,100029,中国;yuesong@ime.ac.cn(SY);zhangzhe1@ime.ac.cn(ZZ);zhangkunpeng@ime.ac.cn(KZ);guohuifang@ime.ac.cn(HG);wangran@ime.ac.cn(RW);doutonghui@ime.ac.cn(TD)2 中国科学院大学微电子学院,北京市玉泉路甲 19 号,100049,中国 3 北京信息科技大学光电测试技术与仪器教育部重点实验室,北京市小营东路 12 号,100192,中国;zdl_photonics@bistu.edu.cn(DZ); zhulianqing@sina.com (LZ) 4 东南大学电子科学与工程系,南京四牌楼 2 号 210018,中国 * 通信地址:h.yang@seu.edu.cn (HY); zz241@ime.ac.cn (ZZ) † 这些作者对本文的贡献相同。

ED 377 078 标题 机构 出版日期 说明... - ERIC
本期环境培训机会汇编包括有关全球非学分、本科至研究生教育机会的信息。研究领域包括:环境教育;环境科学;环境管理;非洲和全球变化;环境健康和毒理学;农林业推广;灌溉项目;国际发展政策;环境法;环境工程;人类生态学;林业;采矿和环境;能源和环境;环境与贸易;环境与发展;水资源管理;濒危物种保护;农业和农村发展。材料分为以下几部分:(1)亚洲和太平洋(5 个条目);(2)非洲(5 个条目);(3)北美(7 个条目);(4)欧洲(24 个条目)。提供地址、日期、电话号码,有时还提供传真号码,以及每个课程、课程或机构的简短描述。(LZ)
ED 377 078 标题 机构 出版日期 说明... - ERIC
本期环境培训机会汇编包括有关全球非学分、本科至研究生教育机会的信息。研究领域包括:环境教育;环境科学;环境管理;非洲和全球变化;环境健康和毒理学;农林业推广;灌溉项目;国际发展政策;环境法;环境工程;人类生态学;林业;采矿和环境;能源和环境;环境与贸易;环境与发展;水资源管理;濒危物种保护;农业和农村发展。材料分为以下几部分:(1)亚洲和太平洋(5 个条目);(2)非洲(5 个条目);(3)北美(7 个条目);(4)欧洲(24 个条目)。提供地址、日期、电话号码,有时还提供传真号码,以及每个课程、课程或机构的简短描述。(LZ)

木薯叶对海南黑山羊抗氧化能力、生长、免疫力及瘤胃微生物代谢的影响
1 中国热带农业科学院热带作物品种资源研究所,农业农村部木薯种质资源保护与利用重点实验室,农业农村部南方作物基因资源与种质创制重点实验室,儋州 571737;limaohn@163.com (ML);lvrenlong@aliyun.com (RL);wenjunou@catas.cn (WO);songbichen@catas.cn (SC) 2 中国热带农业科学院湛江实验站,湛江 524000 3 海南大学热带农林学院,海南省热带特种观赏植物种质资源重点实验室,热带特种林木观赏植物遗传与种质创新教育部重点实验室,儋州 571737; zixuejuan@163.com (XZ); lidongzhang@catas.cn (LZ) * 通讯作者: guanyuhou@126.com (GH); zhouhanlin8@163.com (HZ) † 以上作者对本文贡献相同。

1. 不明飞机t23482及其跟踪
~lGTED ACT!Vf 超过 THO • I • 1\-~S FCLLC\~S: l. ~N Ul\ICENTIFlED 飞机出现在 12C' 千米~ETEPS"Jm~l"1!!:1•~·1J!;tr; 1 2348Z.UND TRA<;.t'ING 该飞机在 40 千米处停止。-·-'-· .. ·••1••• ••• 在 0!H3Z。L2· AN u N 1 D EN -r 1 FI E r 飞机 A::> PE .AR... 11 ~ K r Lo fvl ET EP ~ FA s T o F .JI. A IP AT 0 10 lz lF 无 TRACKit\'hCF 这架飞机于 l0c:: K ILfMF.TFRS NOF.HEASLJOF ... IS· AT .-I 0105Z. ·l.]_. t: . . ·AIRCRt1FT .APFEARED 68 公里'" 东经 C£..) ••. .ATHALTTTUDES CF 8, 000- €,'50!? ~ETE:RS ANO AT SFEEDS Of 110-17e. K.NO S.· T~AC!

氦气和氢:运输的过去和未来?
在1920年代和1930年代的飞艇灾难,包括阿克伦号,梅肯号号航空母舰和LZ 129 Hindenburg等人对飞艇的安全产生了担忧,以及第二次世界大战的出现,固定机翼飞机的发展完成了军事船舶的崩溃。但是,投资到该行业的投资使氦气的提取明显便宜。这导致了它在1939年的呼吸混合物中使用,并在第二次世界大战期间用于生产固定机翼飞机的镁焊接。在1950年代,随着低温技术的发展和氦气中氦气的潜力(现在是量子计算),并且在太空竞赛中使用氦气清除火箭时,人们对潜在的高科技氦的未来有了远见。美国政府在1960年代开始储存大量氦气。在1980年代和90年代氦产业被私有化,美国政府开始出售美国氦气储备。这导致自2004年以来定期发生的氦气供应短缺。

数学专业简讯 - MPA 咨询
请务必阅读您大学的电子邮件和 SAN。按照 SAN 和/或您的顾问的电子邮件中找到的预注册步骤进行操作。步骤 1:对您计划攻读的学位进行学位审核(选择正确的目录)。步骤 2:在春季咨询工作表上列出您计划参加的课程(使用课程编号,而不是唯一编号);在“学生评论”下包含问题或解释。步骤 3:阅读您的顾问在工作表上的回复,因为他们可能会提出建议。如果您想与 Jennifer McHam (jennifer.mcham@austin.utexas.edu) 或 Nathaniel Sulapas (nathaniel.sulapas@austin.utexas.edu) 预约,请直接给他们发送电子邮件。如果 Olivia Biehle 是您的学术顾问,并且您的姓氏以字母 AK 开头,那么 Nathaniel 就是您的临时学术顾问;姓氏以字母 LZ 开头,那么 Jennifer McHam 就是您的临时学术顾问。如果您希望与教职顾问会面,请发送电子邮件至数学、物理和天文学咨询办公室 mpaadv@austin.utexas.edu,通过 Zoom 安排咨询预约。

行政信息 - 北约 ACT
我们已经预订了 MEA 大楼 026 号军事风格客房,可供 65 人入住,从 6 月 10 日星期二到 6 月 12 日星期四,免费入住,不含早餐和毛巾,但包括电视、无线网络和 MEA 健身中心的使用权。请联系 LTC Paul de Ruiter ( paul.deruiter@act.nato.int ) 和 LTC Grigory Medina ( grigory.medina@act.nato.int ) 进行预订(先到先得)。另一个选择是 IBIS Stopera Hotel Valkenburgerstraat 68 1011 LZ Amsterdam(距离 MEA 10 分钟步行路程)。截至 5 月 27 日,我们已预订了 20 间客房,从 6 月 10 日星期二到 6 月 12 日星期四。费用为 101 欧元 - 客房/晚/不含早餐,包括无线网络和增值税。预订属于“NATO”。请联系 IBIS Stopera 酒店 (H3044- re@accor.com) 进行预订。早餐和午餐在 030 号楼,需以现金(欧元)支付。进入海军基地:MEA 是 NLD 海军营房;您需要军人或公民身份证(护照)才能进入。准备:

药物非活性成分对生物靶标的活性
* 通讯作者。bshoichet@gmail.com (BKS);laszlo.urban@novartis.com (LU)。作者贡献:JP、BKS 和 LU 构思了这项研究(KVL 为 LU 提供了意见),收集了其他作者的结果并撰写了手稿;JP 在 JJI 的支持和指导下进行了所有 SEA 计算;LZ 和 KMG 进行了有机阴离子转运蛋白抑制试验并协助进行数据分析;DA、AF 和 SW 构思了体外药理学分析;AF 和 HJ 执行了试验;ELB 设计并执行了 BioMAP 实验;HT 执行了所有聚集试验和分析;X.-PH、SS 和 BLR 进行了 G 蛋白偶联受体相关试验和分析;BB 和 GL 提供了异速建模;DB 设计并执行了药代动力学实验;SNG 提供了关于硫柳汞的临床内容和专家意见。所有作者都为手稿的准备做出了贡献。