XiaoMi-AI文件搜索系统
World File Search SystemmCherry

mCherry 明亮变体的定向进化:......
图片标题 图 S1 C9 和 C12 变体的 EP-PCR 文库。 图 S2 对 mCherry 家族进行诱变努力以实现寿命进化的总结。 图 S3 空间远距离替换对光物理特性的作用。 图 S4 寿命和酵母细胞共同进化轨迹。 图 S5 哺乳动物细胞的细胞亮度。 图 S6 大肠杆菌的光漂白趋势。 图 S7 比较大肠杆菌在激发速率标准化条件下的光稳定性。 图 S8 归一化的吸收和发射光谱。 图 S9 变体的荧光各向异性衰减和旋转时间常数(τ r)。 图 S10 变体的荧光衰减和平均寿命(τ)。 图 S11 变体的荧光量子产率()。 图 S12 mCherry 变体的辐射速率常数分析。图 S13 总非辐射速率常数与能隙的拟合。图 S14 相关可观测量的示意图。图 S15 波数尺度上的吸光度和荧光光谱。图 S16 方程 S2 中的分子和分母函数。补充数据表列表

补充材料
图 S2。反式配对切口方法导致人类 HSPC 中的 HDR 效率低下。(A) 使用 CRISPR/Cas9、单切口或反式配对切口方法在 HEK293T 细胞中将 T2A-mCherry 插入人类 B2M 基因座的靶向策略。使用没有 (pDonor) 或有 2 个靶序列 (TS) (pDonor-Nick 2) 的供体质粒。靶向 B2M 的常见 sgRNA 以红色表示。使用所示方法靶向 B2M 基因座六天后,对 mCherry + HEK293T 细胞的百分比进行 FACS 分析。图表总结了通过 FACS 测量的 mCherry + (HDR) HEK293T 细胞的频率。(B) 使用 CRISPR/Cas9、单切口或反式配对切口方法在人类 HSPC 中将 T2A- mCherry 插入人类 B2M 基因座。使用单链 (ss) AAV 和不含 (scAAV) 或含 2 个 TS 的自互补 (sc) AAV (scAAV-Nick 2 ) 作为供体模板。FACS 分析显示靶向三天后 mCherry + HSPC 的频率。条形图显示 HDR (mCherry + ) 效率。数据显示为四次独立实验的平均值 ± SD。

基于AMA1的下一代质粒,用于增强丝状真菌的异源表达
图1使用下一代AMA1质粒增强了麦克利荧光蛋白的表达。A。分析了MCHERRY表达的AMA1质粒的示意图,其选择标记物具有不同的变体。b荧光曲霉曲霉菌落的荧光照片显示,用Ubi-M-Pyrg和Ubi-Y-Y-Pyrg质粒转化的菌落中荧光增加。来自转化菌落的孢子中麦克利荧光的流式细胞仪分析表明,使用UBI-Y-Y-PYRG质粒实现了最均匀和最高的麦克利信号。在图S1a中,来自不同转化菌落的重复之间的平均荧光和重复的直方图。在液体培养中生长的菌丝体的共聚焦显微镜图像显示,在含有UBI-M-PYRG和UBI-Y-PYRG质粒的菌丝体中的表达增加。ImageJ火灾校准栏代表不同级别的MCHERRY信号。在图中重复S2。e。对等差质粒浓度下不同质粒的转化效率的评估显示,质质质质量降低的质粒的转化效率降低了,将pyRG融合到降解标签。字母表示由ANOVA确定的,并在Tukey后的测试中确定了显着不同的组。F.在选定和非选择条件下固体培养基上菌落生长速率的比较表明,在选择性条件下,携带UBI-M-PYRG和UBI-Y-Y-PYRG质粒的菌株的生长较慢。星号代表pADJ <0.05 <0.05,韦尔奇的t检验表示选择性和非选择性培养基之间的直径差异,用于携带每种质粒的菌株。

使用 AAV 介导的原位基因标记可视化哺乳动物大脑中的 Arc 蛋白动力学和定位
活性调节的细胞骨架相关 (Arc) 蛋白对于突触可塑性和记忆形成至关重要。Arc 基因含有结构 GAG 逆转录转座子序列的残余,它产生的蛋白质可自组装成含有 Arc mRNA 的衣壳状结构。从神经元释放的 Arc 衣壳已被提议作为一种新的 mRNA 传递细胞间机制。尽管如此,仍然缺乏 Arc 在哺乳动物大脑中细胞间运输的证据。为了能够在体内追踪来自单个神经元的 Arc 分子,我们设计了一种腺相关病毒 (AAV) 介导的方法,使用 CRISPR/Cas9 同源独立靶向整合 (HITI) 将荧光报告基因标记到小鼠 Arc 蛋白的 N 端。我们表明,编码 mCherry 的序列可以成功敲入 Arc 开放阅读框的 5′ 端。虽然 Arc 起始密码子周围有 9 个 spCas9 基因编辑位点,但编辑的准确性高度依赖于序列,只有一个靶标导致框内报告基因整合。在海马中诱导长期增强 (LTP) 时,我们观察到 Arc 蛋白的增加与荧光强度和 mCherry 阳性细胞数量的增加高度相关。通过邻近连接分析 (PLA),我们证明 mCherry-Arc 融合蛋白通过与突触后棘中的跨膜蛋白 stargazin 相互作用而保留了 Arc 功能。最后,我们在靠近编辑神经元的 mCherry 阳性棘的 mCherry 阴性周围神经元中记录了 mCherry-Arc 与突触前蛋白 Bassoon 的相互作用。这是第一项为哺乳动物大脑中 Arc 的神经元间体内转移提供支持的研究。
背景方法目标结果cont'...
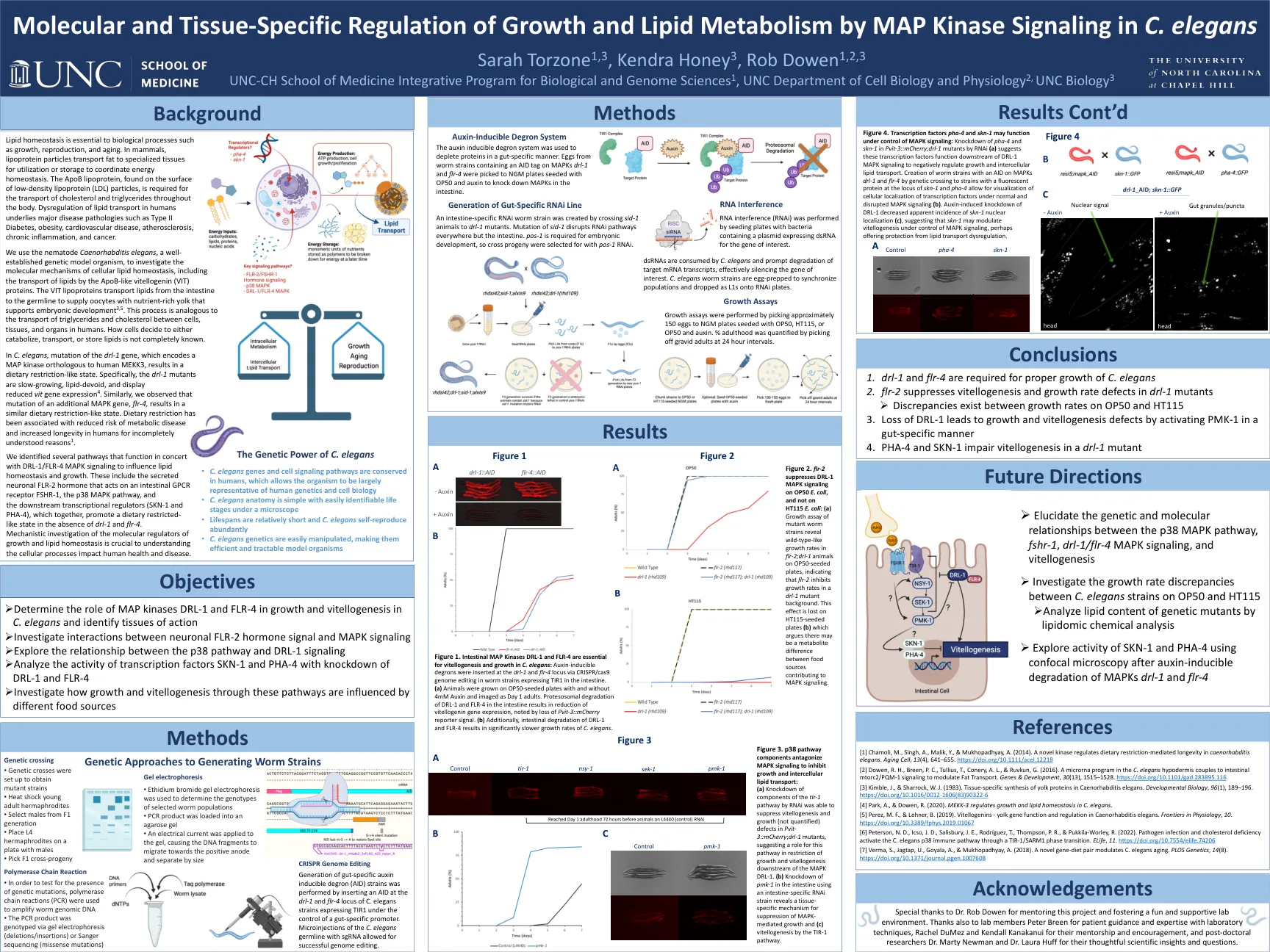
图4。转录因子PHA-4和SKN-1可能在MAPK信号控制下起作用:PVIT-3 :: MCHERRY中PHA-4和SKN-1的敲低; RNAi(a)的DRL-1突变体(a)表明,这些转录因子在DRL-1 MAPK信号的下游作用,以负调节生长和细胞间的Lipid Lipid运输。通过遗传交叉在SKN-1和PHA-4的基因座和PHA-4的菌株中菌株创建蠕虫菌株,并在MAPKS DRL-1和FLR-4上辅助,从而可以在正常和破坏的MAPK信号(B)下可视化转录因子的细胞定位(b)。生长素诱导的DRL-1敲低DRL-1降低了SKN-1核定位(C)的明显发生率(C),这表明SKN-1可能会在控制MAPK信号传导的控制下调节植物生成,也许可以保护脂质转运失调。

Combi‐CRISPR:NHEJ 和 HDR 的结合提供了……
摘要 CRISPR-Cas9 广泛用于小鼠和大鼠的基因靶向。非同源末端连接 (NHEJ) 修复途径在受精卵中占主导地位,可有效诱导插入或缺失 (indel) 突变,从而在靶位点敲除基因,而通过同源定向修复 (HDR) 的基因敲入 (KI) 则难以产生。在本研究中,我们使用双链 DNA (dsDNA) 供体模板与 Cas9 和两个单向导 RNA,一个用于切割目标基因组序列,另一个用于切割 dsDNA 质粒的侧翼基因组区域和一个同源臂,在 G0 幼崽中产生 20-33% 的 KI 效率。 G0 KI 小鼠在一个靶位点携带 NHEJ 依赖的插入/缺失突变,该突变设计在内含子区域,而在另一个外显子位点携带 HDR 依赖的各种供体盒(例如 EGFP 、mCherry 、Cre 和感兴趣的基因)的精确 KI,这些供体盒的长度从 1 到 5 kbp 不等。这些发现表明,这种由 CRISPR-Cas9 系统介导的 NHEJ 和 HDR 组合方法有助于在小鼠和大鼠中高效、精确地 KI 质粒 DNA 盒。

qSanger:通过桑格测序对细菌培养物中的遗传变异进行量化
所有培养生物体中都会自发出现突变和重组等遗传变异。虽然可以通过选择或反选择来识别非中性突变,但在异质群体中识别中性突变通常需要昂贵且耗时的方法,例如定量或液滴聚合酶链反应和高通量测序。在不断变化的环境条件下,中性突变甚至可能成为主导,从而强制进行暂时选择或反选择。我们提出了一种新方法,我们称之为 qSanger,使用来自混合 Sanger 测序读数的对齐电泳图峰的振幅比来量化 DNA。表达增强型绿色荧光蛋白和 mCherry 荧光标记的质粒用于通过定量聚合酶链反应和荧光定量在体外和共转化大肠杆菌中验证 qSanger。我们表明,qSanger 允许从混合 Sanger 测序读数中量化遗传变异,包括单碱基天然多态性或从头突变,与标准方法相比,大大减少了劳动力和成本。

细胞特异性单病毒载体 CRISPR/Cas9 编辑和中枢及外周神经系统中的基因编码工具传递
(AAV)。为了克服这些限制,我们开发了一种替代基因编辑策略,使用单个 AAV 载体和表达 Cre 依赖性 Cas9 的小鼠系,实现整个神经系统内有效的细胞类型特异性编辑。从基因组位点表达 Cre 依赖性 Cas9 提供了空间,可以将用于基因编辑的指导 RNA 与 Cre 依赖性、遗传编码的工具一起包装在一起,以使用单个病毒来操纵、映射或监测神经元。我们用神经科学中的三种常见工具验证了这一策略:ChRonos(一种通道视紫红质),用于使用光遗传学研究突触传递,GCaMP8f 用于使用光度测定法记录 Ca 2+ 瞬变,以及 mCherry 用于追踪轴突投射。我们在多个脑区和细胞类型中测试了这些工具,包括伏隔核中的 GABA 能神经元、从腹侧苍白球投射到外侧缰核的谷氨酸能神经元、腹侧被盖区中的多巴胺能神经元和外周的本体感受神经元。这种灵活的方法可以帮助通过一次病毒注射识别和测试影响突触传递、电路活动或形态的新基因的功能。

摘要PDF海报
摘要:前连合(AC)是一束轴突,它们在嗅觉区域(例如嗅球(OB),前嗅觉核(AON)和梨状皮层(PC)等嗅觉区域之间交流,在嗅觉区域之间进行交流。以前,我们报道说,AC的发展是一个高度调节的过程,涉及渐进式和回归的增长策略,在E17胚胎开发结束时达到对侧。同时,对侧结构中的树博化延迟到产后3-5天。在这里,我们使用与EGFP或MCHERRY转导的腺相关病毒(AAVS)向量,我们在OB,AON和PC中注入了嗅觉区域,以研究穿过AC的对侧神经支配场。我们发现,来自OB的对侧轴突仅穿过AC的前肢,以投射到颗粒细胞层(GCL)中。相比之下,轴突源自前PC项目,进入对侧OB,AON和PC。这些轴突不仅将其释放到GCL中,还可以伸入二尖瓣和外部丛状层,以及前PC层1B。,我们通过AC的后肢专门观察到后PC项目,专门于对侧PC,从1B层进行了根本性的塑造。内一核核仅通过AC的后肢向后PC进行。共同展示了嗅觉结构中对侧树博化的详细图,这对于理解脑半球之间嗅觉信息的处理至关重要。

TET调节的表达和光学清除,用于遗传编码的嵌合DCAS9/荧光蛋白探针的体内可视化
摘要:Cas9(DCAS9)核酸内切酶的催化无效突变体具有多种生物医学应用,最有用的是转录的激活/抑制。dcas9家族成员也正在成为潜在的实验工具,用于在独立活细胞和完整组织的水平上进行基因映射。我们对CAS9介导的核室可视化的一组工具进行了初步测试。我们研究了doxycycline(DOX) - 可诱导(TET-ON)的细胞内分布,这些构建体的构造中编码DCAS9直系同源物(ST)(ST)和脑膜炎N.脑膜炎(NM)与EGFP和MCHERRY FOLORESCENT蛋白(FP)融合的人类A549细胞。我们还研究了这些嵌合荧光构建体的时间依赖性表达(DCAS9-FP)在活细胞中诱导中的诱导中,并将其与实验性DCAS9-FP表达的时间过程进行了比较灌注。在诱导后24小时内,肿瘤异种移植物发生了麦克利 - 奇氏菌表达的体内诱导,并通过使用皮肤的光学清除(OC)来可视化。OC通过局部应用Gadobutrol启用了肿瘤异种移植物中FP表达的高对比度成像,因为红色和绿色通道的FI增加了1.1-1.2倍。