XiaoMi-AI文件搜索系统
World File Search Systemmutation

直接和间接突变分析I:PCR
•细胞裂解和核酸酶灭活后,可以通过过滤或离心轻松从裂解物中除去细胞碎片。•接下来是去除蛋白质和RNA。•通常,通过用蛋白水解酶(例如蛋白酶K)消化大多数蛋白质,该蛋白酶K具有活性,而蛋白酶K具有广泛的天然蛋白质。•大部分RNA被切开时释放的RNass切割。•DNA提取物中RNA的存在不是主要问题,因为这不会干扰PCR或限制消化。•在某些情况下,重要的是要分离不含RNA的DNA,以便能够使用分光光度计准确地量化DNA产量。•浓缩DNA最广泛使用的方法是用乙醇沉淀。•用酒精(通常是乙醇或异丙醇)沉淀DNA。由于DNA不溶于这些酒精,因此会聚集在一起,在离心时给出颗粒。此步骤还去除了酒精可溶性盐


D1.3 突变和基因编辑 - 生物信件
• 前导序列:位于 CRISPR 基因座一端的非编码序列(长度为 80-500 个核苷酸),有助于启动 RNA 转录并整合新的入侵者基因组(间隔物)。 • 间隔物:与入侵者(即病毒物质)相匹配的短而独特的 DNA 序列,本质上是原核生物免疫系统的记忆。 • 重复序列:分隔每个间隔物的短而相同的 DNA 序列。它们有规律地间隔开来,通常是回文结构(从 5' 和 3' 方向对称),这就是 CRISPR 这个首字母缩略词的由来“成簇的、有规律间隔的、短回文重复序列”。 位于 CRISPR 阵列附近的是 cas 基因,它们是编码区,用于编码蛋白质复合物的合成,如 Cas 蛋白(因此得名 CRISPR-Cas 系统),Cas 蛋白是一种能够消化 DNA 的核酸酶。当病毒入侵原核生物时,与病毒遗传物质相匹配的 CRISPR 阵列会转录成单个向导 RNA (sgRNA),该 RNA 会与 Cas 蛋白结合并引导其朝向病毒的遗传物质。当 sgRNA 检测到匹配的病毒 DNA 时,Cas 蛋白会裂解/切割 DNA,从而有效地阻止病毒感染。

用 QISKit 编写的量子程序的变异测试
量子计算机的快速普及势必会打破数十年来的若干计算限制,但它也势必会在软件测试等领域带来重大挑战。测试是指在复制真实场景的体外环境中执行软件以确定其正确行为 [3]。尽管如此,在经典计算领域,测试已经得到了广泛的研究,并且已经提出了多种方法和工具 [1],但此类量子程序 (QP) 方法仍处于起步阶段 [10]。值得注意的是:(i) QP 比经典程序难开发得多,因此,大多数熟悉经典世界的程序员更容易在违反直觉的量子编程中犯错误 [6];(ii) QP 必然是概率性的,不可能在不中断执行或不损害其结果的情况下进行检查 [7]。因此,确保 QP 的正确实现在量子计算领域更具挑战性 [4]。

EnGen 突变检测试剂盒 E3321 手册
EnGen 突变检测试剂盒提供用于检测靶向基因组编辑事件的试剂。第一步,使用 Q5 Hot Start High-Fidelity 2X Master Mix 扩增基因组被靶向的细胞(即 CRISPR/Cas9、TALEN、锌指核酸酶)的目标区域。变性和重新退火后,当扩增子池中存在插入和缺失 (indel) 突变时,会形成异源双链。第二步,退火的 PCR 产物用 EnGen T7 核酸内切酶 I 消化,这是一种结构特异性酶,可识别大于 1 个碱基的错配。当存在错配时,DNA 的两条链都会被切断,从而形成较小的片段。对所得片段的分析可以估计基因组编辑实验的效率。
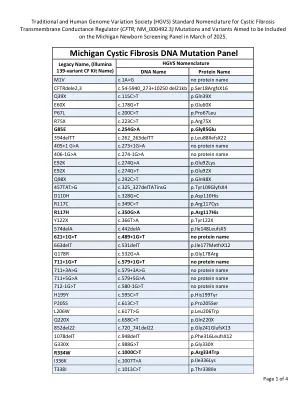
密歇根囊性纤维化 DNA 突变检测小组
R347P c.1040G>C p.Arg347Pro R347H c.1040G>A p.Arg347His R352Q c.1055G>A p.Arg352Gln 1213delT c.1081delT p.Trp361GlyfsX8 1248+1G>A c.1116+1G>A 无蛋白质名称 1259insA c.1127_1128insA p.Gln378AlafsX4 W401X(c.1202G>A) c.1202G>A p.Trp401X W401X(c.1203G>A) c.1203G>A p.Trp401X 1341+1G>A c.1209+1G>A 无蛋白质名称 5T* c.1210-12T[5] 无蛋白质名称 7T* c.1210-12T[7] 无蛋白质名称 9T* c.1210-12T[9] 无蛋白质名称 1461ins4 c.1329_1330insAGAT p.Ile444ArgfsX3 A455E c.1364C>A p.Ala455Glu 1525-1G>A c.1393-1G>A 无蛋白质名称 S466X(C>A) c.1397C>A p.Ser466X S466X(C>G) c.1397C>G p.Ser466X L467P c.1400T>C p.Leu467Pro 1548delG c.1418delG p.Gly473GlufsX54 S489X c.1466C>A p.Ser489X S492F c.1475C>T p.Ser492Phe Q493X c.1477C>T p.Gln493X I506V** c.1516A>G p.Ile506Val

Engen突变检测试剂盒E3321手册
ENGEN突变检测试剂盒提供了用于检测目标基因组编辑事件的试剂。在第一步中,使用Q5热启动High-Fidelity 2X Master Mix放大了来自基因组的靶向区域(即CRISPR/CAS9,TALES,锌指核酸酶)。在变性和重新进行重新进行后,当插入和缺失(Indels)中存在于扩增子池中时,就会形成异质化合物。在第二步中,将退火的PCR产物用Engen T7核酸内切酶I消化,这是一种特定于结构的酶,将识别大于1碱基的不匹配。存在不匹配时切割DNA的两个链,从而导致形成较小的片段。对所得片段的分析提供了基因组编辑实验效率的估计。

poziotinib用于HER2外显子20插入突变
* HER2外显子20插入突变的患者ORR:总反应率; ICR:独立中央评论; DOR:响应持续时间; EGFR:表皮生长因子受体; HER2:人表皮生长因子受体2

循环肿瘤 DNA 突变分析及配对...
摘要:循环肿瘤DNA(ctDNA)是一种常规的液体活检样本,可通过采集来动态监测疾病状态。然而,其潜在的临床价值以及与腹水样本或肿瘤活检的一致性对于卵巢癌患者还有待进一步评估。因此,本研究比较了ctDNA、配对肿瘤组织和腹水样本之间的突变谱,以探讨它们在卵巢癌中的可能临床价值。使用靶向下一代测序筛查卵巢癌患者18个外周血样本、6个配对腹水样本和8个配对肿瘤组织中的突变。使用公共数据库进行功能分析。使用WebGestalt进行基因本体论和通路富集分析。使用cBioPortal for Cancer Genomics评估治疗靶点。使用Chilibot和检索相互作用基因/蛋白质的搜索工具来获取关键基因及其功能相互作用。使用维恩图对三类样本进行比较分析。在ctDNA样本中共发现104个癌症相关突变基因,在肿瘤组织中共发现95个基因,在腹水样本中共发现44个基因。通过Chilibot分析获得了一个涵盖10个基因的簇,即NOTCH2、NOTCH3、赖氨酸甲基转移酶2A、PTEN、雄激素受体、DNA活化蛋白激酶催化亚基、肝细胞核因子1同源框A、SRC、胰岛素受体底物2和SRY盒转录因子10。该基因组可能具有监测卵巢癌转移和确定治疗靶点的潜力

优化ABE和GRNA以纠正G6PC-R83C突变
糖原储存疾病IA型(GSDIA)是由G6PC基因突变引起的常染色体隐性疾病,它破坏了葡萄糖稳态中的关键酶G6Pase 1。GSDIA患者患有低血糖,肝脏和肾脏的糖原和脂肪的积累,导致肝肿大和肾肿大。无法治愈。急性致命的低血糖,但肾脏疾病和肝细胞癌的长期并发症并未解决。与GSDIA相关的两个最普遍的G6PC突变是R83C和Q347X,均包含单个G> A的过渡突变。腺嘌呤碱基编辑器(ABES)可以使用基因组DNA中A•T到G•C的编程转换,并且原理可以用来精确纠正这些突变。在这里,我们设计了新颖的腺嘌呤基础编辑器(ABE)变体,以验证GSD1A的临床模型。