XiaoMi-AI文件搜索系统
World File Search Systemnhi


2024-26 年 NHI 战略
我们积极让各级会员参与进来,齐心协力推动爱尔兰成为老年人护理领域的典范。通过加强参与结构和赋予会员发言权,我们确保他们的经历塑造未来服务的发展。我们的会员为居民提供全天候专业护理,我们将继续支持他们的需求,同时寻找新的方式来建设我们的社区并扩大我们的服务范围。

NHI 集团 - 2023 年 12 月 31 日年度财务报告
雀巢控股公司(“NHI”)(以下与其子公司一起称为“NHI 集团”)在美国特拉华州注册成立,是 NIMCO US, Inc. 的全资子公司,而 NIMCO US, Inc. 是雀巢公司的间接全资子公司,雀巢公司在瑞士注册成立,是雀巢集团的控股公司(以下称为“雀巢集团”)。NHI 是雀巢公司在美国的主要运营子公司的控股公司,其中包括雀巢美国公司、雀巢普瑞纳宠物护理公司和嘉宝产品公司。NHI 集团主要从事食品、宠物护理产品、优质饮用水、饮料以及营养和健康科学产品的制造和销售。这些业务的收入来自美国和国际市场。

濒危资源认证计划指南
自然遗产清单(NHI)计划:由NATURESERVE建立并目前由Natureserve协调的国际NHI计划的一部分。所有NHI计划都使用标准方法来收集,表征和管理数据,从而可以在各种尺度上组合数据以解决本地,州,区域和国家问题。NHI计划着重于定位和记录稀有物种和自然社区的发生,包括州和联邦的濒临灭绝和受威胁的物种。物种和自然社区,可以在NHI工作列表中找到。提出的濒危资源(ER)评论:拟议的ER评论详细介绍了稀有物种,高质量的自然社区以及其他可能受到特定拟议项目影响的濒危资源,以及避免对这些资源产生负面影响的建议。所有提议的ER审查均根据文件“进行拟议的濒危资源(ER)评论:逐步认证ER审稿人的逐步指南”的要求起草。

南非卫生技术部门的概述 荷兰海上风创新指南2024 挪威拔掉电池价值链 澳大利亚关键的矿物机会| 2023 Horizon Europe Klankbordgroep群集4-数字行业... 荷兰氢经济的解决方案。pdf
但是,南非的医疗保健部门目前处于易流量状态,火车中的几个过程可能会导致该行业的结构性变化。最重要的是,如果实施,政府提议的国家健康保险(NHI)计划将从根本上改变公共部门和私营部门的运作方式。在此阶段,NHI的外观细节以及计划是否会付诸实践,仍然存在很多不确定性。但是,作为中期战略的一部分,所有医疗保健产品和服务的提供者都应开始考虑如何将自己定位为NHI基金的供应商。nhi机会将由能够扩大规模以应对实现国家足迹的挑战(并且能够证明可以扩大规模的挑战)抓住机会。

s12913-021-07360-w.pdf
摘要背景:附加适应症的发展战略与药品价格修订之间的关系尚未得到深入研究。在此,我们调查了日本批准的抗癌药物的价格修订。方法:该研究基于日本 2009 年 1 月至 2020 年 3 月期间批准的抗癌药物的已发布信息。我们调查了抗癌药物的药理和监管特征与日本国民健康保险 (NHI) 价格修订的发生/不发生之间的关系。结果:在调查期间,81 种新的抗癌药物被列入 NHI 价格清单。2020 年 4 月 1 日,23 种抗癌药物的价格已从初始定价修订,21 种药物(91.3%)的价格降低。几个参数显示了药物特性和 NHI 价格修订之间的关系。附加适应症的实现和化合物类型被确定为这些关系的解释因素。定义了附加适应症概况以评估附加适应症实现方法与价格修订之间的关系。追加适应症类型为“扩大”时,药品获得 NHI 价格修订的比例最高( P <0.001)。结论:NHI 价格修订与追加适应症的实现和化合物类型显著相关。发现追加适应症的策略会影响 NHI 价格修订的发生/不发生。关键词:抗癌药物,肿瘤学,药品价格修订,追加适应症,日本,国民健康保险药品价格
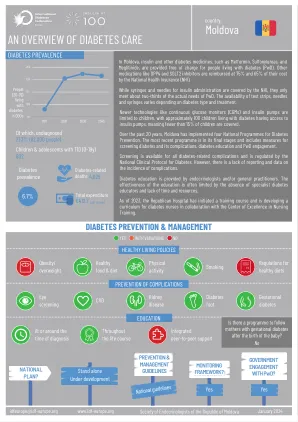
摩尔多瓦(004)-IDF Europe
在摩尔多瓦,胰岛素和其他糖尿病药物中,例如二甲双胍,磺氟脲和丙列他汀,为患有糖尿病患者(PWD)的人免费提供。由国家健康保险(NHI)(NHI)以75%和65%的成本偿还了其他药物,例如DPP4和SGLT2抑制剂的75%和65%。

医院限制清单 - 脊髓灰质炎疫苗
病房: ................................................................................................................ NHI: ......................................................................................................................

表格 6-k 美国证券交易委员会 - 野村
(1)最近五个财务年度的选定合并财务数据。截至 3 月 31 日的年度 2018 年 2019 年 2020 年 2021 年 2022 年 总收入(百万日元) 1,972,158 1,835,118 1,952,482 1,617,235 1,593,999 净收入(百万日元) 1,496,969 1,116,770 1,287,829 1,401,872 1,363,890 税前收入(亏损)(百万日元) 328,158 (37,701) 248,261 230,671 226,623 归属于野村控股株式会社(“NHI”)股东的净收入(亏损)(百万日元) 219,343 (100,442) 216,998 153,116 142,996 归属于本公司股东的综合损益(损失)(百万日元) 126,335 (70,136) 219,943 141,077 309,113 股东权益总额(百万日元) 2,799,824 2,680,793 2,731,264 2,756,451 2,972,803 资产总额(百万日元) 40,343,947 40,969,439 43,999,815 42,516,480 43,412,156 每股股东权益(日元) 810.31 794.69 873.26 879.79 965.80 归属于本公司普通股股东的每股净利润(亏损)—基本(日元) 63.13 (29.90) 67.76 50.11 46.68 归属于本公司普通股股东的每股净利润(亏损)—稀释(日元) 61.88 (29.92) 66.20 48.63 45.23 本公司股东权益占总资产的比例(%) 6.8 6.4 6.0 6.3 6.7 股东权益回报率(%) 7.92 (3.73) 8.21 5.73 5.10 市盈率(倍) 9.75 — 6.76 11.60 11.04 营业活动产生的现金流量(百万日元) (445,690) (361,165) (15,943) 665,770 (1,368,710) 投资活动产生的现金流量(百万日元) (56,172) (112,503) 216,336 (139,026) (45,301) 融资活动产生的现金流量(百万日元) 373,168 761,191 332,062 (269,927) 1,070,715 年末现金、现金等价物、限制性现金及限制性现金等价物(百万日元) 2,354,868 2,687,132 3,192,310 3,510,011 3,316,408 员工人数 28,048 27,864 26,629 26,402 26,585 [临时员工平均人数,不包括上述人数] [4,671] [4,492] [4,313] [4,224] [4,339] 1 野村控股公司及其合并子公司(“野村”)的选定财务数据按照美国公认会计原则(“美国GAAP”)列示。2 每股股东权益、NHI 股东权益总额占总资产的百分比、股东权益回报率均使用 NHI 股东权益总额计算。3 应税交易的消费税和地方消费税按免税法核算。4 员工人数中包含某些合同工。6 由于净亏损,未列明截至 2019 年 3 月 31 日止年度的市盈率(倍数)。25 由于会计准则更新 2014-09“来自客户合同的收入”以及野村于 2018 年 4 月 1 日采用的会计政策变化,对之前报告的金额进行了某些重新分类,以符合本年度的列报。

COVID-19 疫苗接种常见问题
● 当父母或监护人预约接种 COVID-19 疫苗时,他们还可以为他们家庭中任何 12-15 岁的孩子预约接种疫苗。 ● 如果没有预约,我们不能接待任何人。 ● 可以通过访问 www.BookMyVaccine.nz 或致电 0800 28 29 26(每周 7 天,上午 8 点至晚上 8 点)进行预约。 我不是新西兰公民或居民,可以接种疫苗吗? 是,无论公民身份或签证身份如何,新西兰的每个人都可以免费接种 COVID-19 疫苗。 我没有在新西兰的医生或全科医生处登记,也没有国家健康指数 (NHI) 编号,我可以接种疫苗吗? 是,你可以接种疫苗。拥有健康编号(国家健康指数或 NHI)不是预约或接种疫苗的必要条件。 新西兰有哪些疫苗?如何使用?