XiaoMi-AI文件搜索系统
World File Search Systempolymerase

聚合酶链反应(PCR)
•离心机/微型/微型旋转 - 用于确保样品/试剂位于管/板的底部,在某些过程中,用于分离混合物的成分成分。•机器人 - 用于自动化某些技术。•涡流 - 用于混合试剂或样品。•热块 - 用于在特定温度下保持样品/反应。•移液器 - 用于转移液体的设定体积。•溢出套件 - 用于清理较大的试剂溢出。•手套 - 用于保护用户和样品。•实验室外套 - 用于保护用户和样品;不使用时需要挂起实验室外套,并经常洗涤以确保它们干净。

TAQ DNA聚合酶(1000)
AIIC-澳大利亚工业化学品库存; ASTM-美国材料测试学会; BW-体重; CERCLA-全面的环境响应,赔偿和责任法; CMR-致癌,诱变剂或生殖有毒物; DIN-德国标准化研究所的标准; DOT-运输部; DSL-国内物质清单(加拿大); ECX-与X%响应相关的浓度; EHS-极其危险的物质; ELX-与X%响应相关的负载率; EMS-紧急时间表; ENC-现有的和新的化学物质(日本); ERCX-与X%增长率响应相关的浓度; ERG-紧急响应指南; GHS-全球统一的系统; GLP-良好的实验室实践; HMIS-危险材料识别系统; IARC-国际癌症研究机构; IATA-国际航空运输协会; IBC-国际批量携带危险化学物质的船舶建筑和设备的国际守则; IC50-最大抑制浓度;国际民航组织 - 国际民航组织; IECSC-中国现有化学物质的清单; IMDG-国际海事危险货物; IMO-国际海事组织; ISHL-工业安全与卫生法(日本); ISO-国际标准化组织; Keci-韩国现有化学品库存; LC50-致命浓度占测试人群的50%; LD50-致死剂量至测试人群的50%(中位致死剂量); Marpol-预防船舶污染的国际公约; MSHA-矿山安全与健康管理; N.O.S.- 不否则

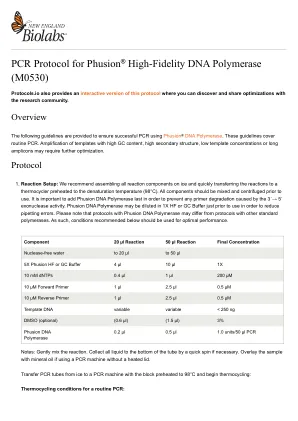
Taq DNA 聚合酶的 PCR 方案
聚合酶链式反应 (PCR) 是一种功能强大且灵敏的 DNA 扩增技术 (1)。Taq DNA 聚合酶是一种广泛用于 PCR 的酶 (2)。以下指南旨在确保使用 NEB 的 Taq DNA 进行 PCR 成功

Taq DNA 聚合酶的 PCR 方案
聚合酶链式反应 (PCR) 是一种功能强大且灵敏的 DNA 扩增技术 (1)。Taq DNA 聚合酶是一种广泛用于 PCR 的酶 (2)。以下指南旨在确保使用 NEB 的 Taq DNA 进行 PCR 成功
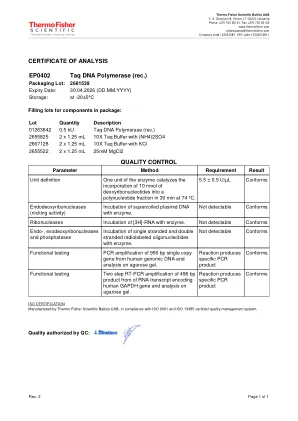
EP0402 TAQ DNA聚合酶(rec。) Invitrogen转染产品 Lentipool V2人CRISPR库 - 用户指南 实现特定于目标效率-truecut-hifi-cas9-
批量描述01263642 0.5 KU TAQ DNA聚合酶(rec。)2655825 2 x 1.25 ml 10x TAQ缓冲液(NH4)2SO4 2667128 2 x 1.25 ml 10x TAQ缓冲液与KCl 2655522 2 x 1.25 ml 25mm 25mm 25mm mgcl2

Pfu DNA 聚合酶综合综述
Pfu DNA 聚合酶是一种源自超嗜热古菌 Pyrococcus furiosus 的耐热酶,因其高保真度和强大的加工性而广受认可。它的 3'-5' 核酸外切酶活性使其成为正确扩增短链和复杂 DNA 链不可或缺的酶。Pfu DNA 聚合酶的这些生化特性促进了其提取和生产方法的重大进步。本综述涵盖了一些传统的纯化方法,包括蛋白质纯化和亲和层析,以及重组基因表达、自动化生产系统和基于膜的技术的最新进展。最近开发了新的酶工程方法,例如 CRISPR-Cas9 介导的基因优化,这提高了提取效率的标准以满足新兴需求。曾经具有挑战性的 Pfu DNA 聚合酶生产已通过在实验室和商业规模的大肠杆菌中重组表达得到了显着简化。涉及 IPTG 浓度和响应面方法的优化技术已将产量提高了 30%。自诱导意味着可以实现更高的生物量输出。如今,Pfu DNA 聚合酶的应用范围从标准 PCR 到分子生物学、法医分析、临床微生物学和生物技术领域的高级临床诊断。

Primestar®MAXDNA聚合酶Ver.2
请勿使用。 - 在未经Takara Bio批准的情况下,转售或转移产品,更改产品进行转售或转移或将其用于制造商业产品。 - 请访问我们的网站以获取有关许可和其他信息的最新信息。 - 此传单中列出的公司名称和产品名称是其各自所有者的公司名称或注册或未注册的商标,这些都是其各自所有者的财产。 - 截至2025年2月1日,此传单中列出的价格是建议的零售价。价格不包括消费税。创建于2025年2月G

Neo Gold TAQ DNA聚合酶
neo Gold Taq DNA聚合酶是一种预混合的,可用的2x溶液,其中包含Anɵ体介导的热启动的NeotAQ DNA聚合酶,DNTPS,MGCL2,增强剂和稳定剂,用于有效的放大器。neo Gold TAQ DNA聚合酶可防止非特异性产物构造,并允许聚合酶链重新组合(PCR)在环境温度下进行,这是使用Anɵ体体修饰聚合酶实现的。
