XiaoMi-AI文件搜索系统
World File Search Systempolymerase

最高nzyprood DNA聚合酶| abo
PCR引物的长度通常在15-30个基础范围内,旨在侧面感兴趣的区域。引物应包含40–60%的GC,并避免可能产生内部二级结构的序列。底漆的3端不应是互补的,以避免产生底漆二聚体。引物二聚体不必要地从反应中去除底漆,并导致与所需反应竞争的不良聚合酶反应。避免在底漆的3末端连续三个G或C核苷酸,因为这可能导致非特异性底漆退火增加了不良反应产物的合成。理想情况下,两个引物的温度应几乎相同(T m),以便在大致相同的温度下使用变性模板DNA退火。

takara ex Premier™DNA聚合酶
通知购买者我们的产品仅用于研究用途。它们不得用于任何其他目的,包括但不限于用于人类,治疗或诊断的使用或商业用途。我们的产品不得转移到第三方,转售,用于转售或用于制造商业产品或未经我们事先书面批准为第三方提供服务。您对此产品的使用也符合该产品网页上takarabio.com上所述的任何适用许可要求。您有责任审查,理解并遵守此类陈述所施加的任何限制。

渗入聚合酶历史 - 创新的力量
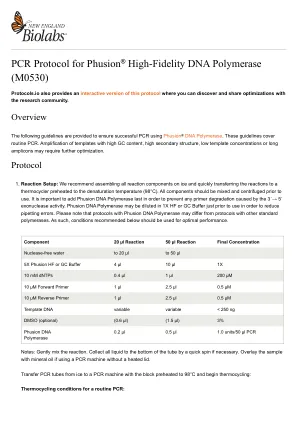
Thermo Scientific™Phusion™高保真DNA聚合酶启用了高性能PCR。自推出以来,Thermo Scientific™Phusion™产品已不断改进,以应对更多的应用和挑战。最新的添加(Thermo Scientific™Phusion™加上DNA聚合酶)适用于许多应用,因为它的高忠诚度,鲁棒性,抑制剂耐受性和通用引物退火。

PrimeSTAR™ MAX Ver.2 DNA 聚合酶
促销代码:TKPRIMESTARV2Q324。促销有效期至 2024 年 10 月 31 日 不适用于预先协商的价格和运费,包括报价、长期订单协议和采购目录协议。所有价格均不含商品及服务税。下订单时请参考促销代码。运费适用。价格如有变动,恕不另行通知。请联系 Scientifix 了解更多产品信息、定价或索取报价。如需客户服务,请致电 1800 007 900 或发送电子邮件至 info@scientifix.com.au。更多信息请访问 www.scientifix.com.au

Primestar®MAXDNA聚合酶Ver.2
请注意,此产品仅用于研究用途。它不打算用于人类或动物的治疗或诊断程序。另外,请勿将此产品用作食品,化妆品或家居用品等。takara产品不得转售或转让,修改用于转售或转让,或无需未经Takara Bio Inc.的书面批准而用于制造商业产品。如果您需要其他使用许可证,请通过我们的网站www.takarabio.com与我们联系。您对此产品的使用也符合产品网页上所述的任何适用许可要求。您有责任审查,理解并遵守此类陈述所施加的任何限制。所有商标都是其各自所有者的财产。某些商标可能不会在所有司法管辖区注册。

Primestar®MAXDNA聚合酶Ver.2
请注意,此产品仅用于研究用途。它不打算用于人类或动物的治疗或诊断程序。另外,请勿将此产品用作食品,化妆品或家居用品等。takara产品不得转售或转让,修改用于转售或转让,或无需未经Takara Bio Inc.的书面批准而用于制造商业产品。如果您需要其他使用许可证,请通过我们的网站www.takarabio.com与我们联系。您对此产品的使用也符合产品网页上所述的任何适用许可要求。您有责任审查,理解并遵守此类陈述所施加的任何限制。所有商标都是其各自所有者的财产。某些商标可能不会在所有司法管辖区注册。

聚合酶参与线粒体DNA维持...
摘要:由活性氧(ROS)触发损坏的线粒体DNA(mtDNA),迄今为止了解到MTDNA维持的过程鲜为人知,这些过程与DNA修复,DNA降解和DNA复制之间的复杂相互作用协调。这项研究旨在通过应用特殊的远程PCR,反映MTDNA完整性,以识别MTDNA维持中涉及的蛋白质。在强制氧化磷酸化的条件下,对基于文献的候选者进行了siRNA筛查,揭示了聚合酶的功能群及其中的聚合酶ζ(POLZ)作为最高命中率。因此,Polz敲低引起mtDNA积累,这需要基础切除修复(BER)核酸酶APE1的活性,然后由单细胞线粒体原位杂交方案(MTRIP)确定的代偿性mtDNA复制。线粒体中的活性氧(ROS)揭示了Polz在次要弧区域形成典型缺失的额外的,无ROS的参与。与证明Polz在线粒体中定位的数据一起,我们建议Polz在mtDNA周转率中起着重要作用,尤其是在氧化应激条件下。

聚合酶链式反应 (PCR) #1 模块小时数
Boster 抗体和 ELISA 专家。(nd)PCR 原理。Boster 抗体和 ELISA 专家。https://www.bosterbio.com/protocol-and-troubleshooting/pcr-principle New England BioLabs Inc. (2021)。使用标准 Taq 缓冲液 (M0273) 的 Taq DNA 聚合酶 PCR 方案。New England BioLabs Inc. https://www.neb.com/protocols/0001/01/01/taq-dna-polymerase-with-standard -taq-buffer-m0273 Pilotte, N. (2013 年 11 月 5 日) 如何计算 PCR 反应所需的引物量?Research Gate。https://www.researchgate.net/post/How_does _one_calculate_the_required_amount_of_primers_required_for_PCR_reaction Promega。(2018 年 4 月)。 GoTaq® DNA 聚合酶 (M300) 协议。Promega。

Titanium®TAQDNA聚合酶分析证书
我们的产品仅用于研究用途。它们不得用于任何其他目的,包括但不限于用于人类,治疗或诊断的使用或商业用途。我们的产品不得转移到第三方,转售,用于转售或用于制造商业产品或未经我们事先书面批准为第三方提供服务。