XiaoMi-AI文件搜索系统
World File Search Systempotent


微核不是 cGAS/STING 通路的强效诱导剂
微核 (MN) 与先天免疫反应有关。MN 膜的突然破裂会导致 cGAS 积聚,从而可能激活 STING 和下游干扰素反应基因。然而,缺乏将 MN 和 cGAS 激活联系起来的直接证据。我们开发了 FuVis2 报告系统,该系统能够可视化携带单个姊妹染色单体融合的细胞核,从而可视化 MN。使用配备 cGAS 和 STING 报告基因的 FuVis2 报告基因,我们严格评估了 MN 在单个活细胞中激活 cGAS 的效力。我们的研究结果表明,在间期,cGAS 定位到膜破裂的 MN 的情况很少,cGAS 主要在有丝分裂期间捕获 MN 并保持与细胞浆染色质结合。我们发现,有丝分裂期间的 cGAS 积累既不会在随后的间期激活 STING,也不会触发干扰素反应。伽马射线照射可独立于微核形成和 cGAS 定位到微核来激活 STING。这些结果表明,细胞质微核中的 cGAS 积累并不是其激活的有力指标,微核不是 cGAS/STING 通路的主要触发因素。

一种有效的 HIV-1 治疗疫苗正在临床试验中
国家。1 根据联合国艾滋病规划署的估计,自疫情开始以来,HIV-1 已影响了 8420 万人,2021 年新增感染病例约 150 万例。据估计,2021 年全球有 3840 万人感染 HIV-1,2870 万人正在接受抗逆转录病毒疗法 (ART) ( https://www.unaids.org/en/resources/fact-sheet )。高效抗逆转录病毒疗法 (HAART) 已被证明是有效预防 HIV-1 相关临床进展的标准治疗方法。2 HAART 成功降低了病毒在靶 T 细胞中的复制和 HIV-1 传播风险,但不能根除感染并抑制血浆病毒载量 (pVL)。 3 此外,急性和慢性药物毒性、耐药菌株的产生以及终生抗逆转录病毒治疗的高成本是抗逆转录病毒药物最重要的问题。 4,5 因此,开发一种廉价的预防性和/或治疗性 HIV-1 疫苗已被提出作为迫切需要。预防性疫苗是阻止 HIV-1 大流行最有希望的解决方案,但它们在 II 期和 III 期人体临床试验中屡屡失败。 6,7 到目前为止,最有效的预防性疫苗是 RV144(ALVAC HIV-1(vCP1521)病毒载体初免/AIDSVAX B/E gp120 蛋白加强)在 III 期临床试验中有效率为 31%,且持久性有限。 8 HIV-1 疫苗开发的生物学障碍源于病毒特性,例如病毒复制过程中的高突变和重组率、遗传多变性和病毒的细胞相关传播。 9、10此外,缺乏适当的艾滋病动物模型和有关 HIV-1 保护的免疫相关性信息有限,都是疫苗研发面临的科学挑战。11、12 目前尚无批准用于临床的预防性 HIV-1 疫苗,新感染 HIV-1 的人数持续增加。因此,开发一种有效的治疗性疫苗作为解决 HIV-1 持续性和治愈感染患者的策略将是一个宝贵的进步。13 此外,由于缺乏针对 HIV-1 感染的明确治疗方法,治疗性疫苗作为 HAART 替代方案的重要性更加凸显。14 到目前为止,由于 HIV-1 基因多样性及其对免疫系统的逃避,不同的治疗性疫苗都无法成功根除病毒。因此,新型治疗性疫苗候选物正在开发中,作为 HIV-1 感染的治疗策略。新的研究经常评估候选疫苗(例如,保守的多表位疫苗构建体)与其他疗法和/或新配方和免疫方案联合使用的有效性。本综述简要介绍了这些保守的多表位构建体的设计,并概述了这些候选疫苗在最近的临床管线中的结果。
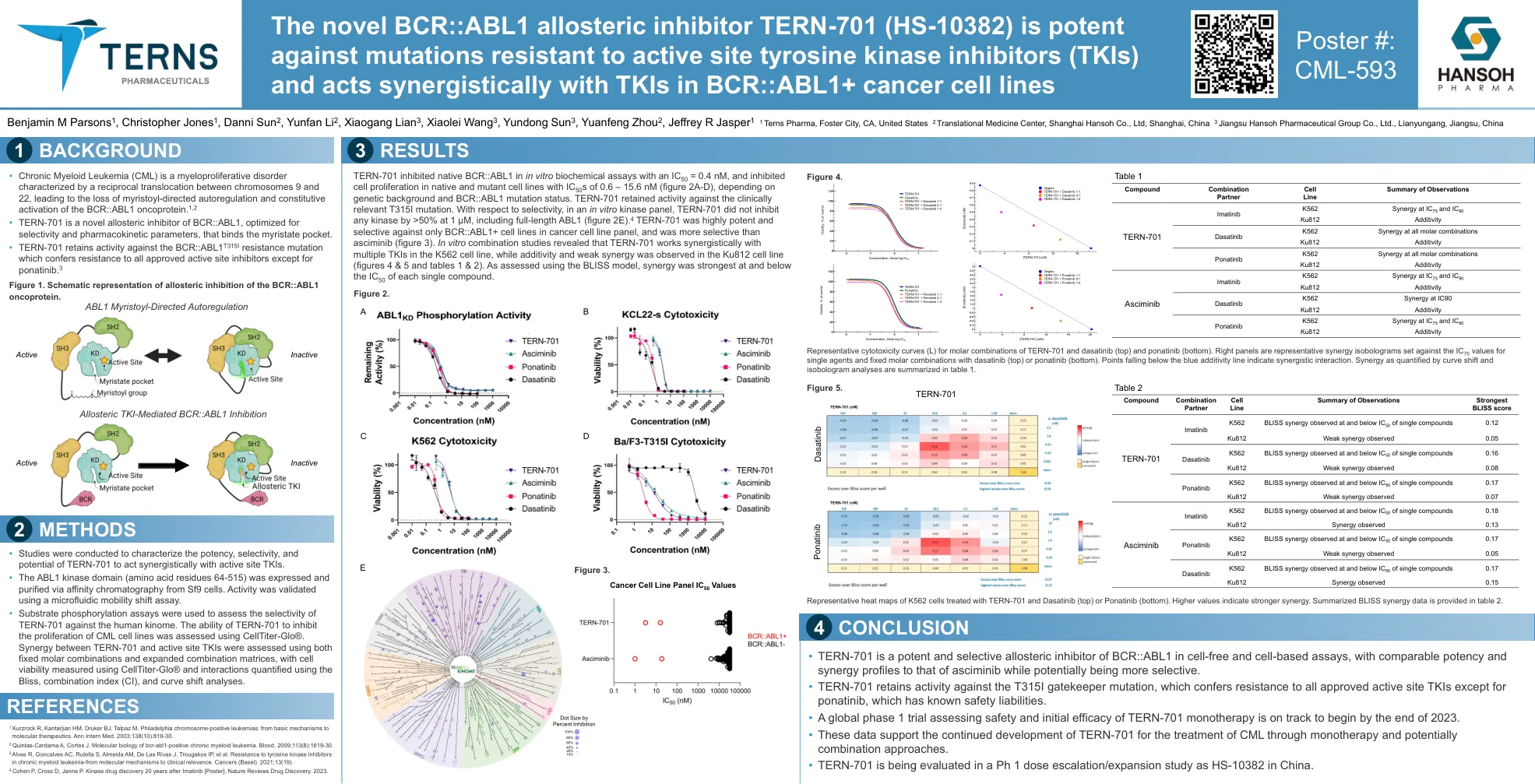
ABL1变构抑制剂Tern-701(HS-10382)有效
•进行研究以表征Tern-701与主动位点TKIS协同作用的效力,选择性和潜力。•ABL1激酶结构域(氨基酸残基64-515)通过SF9细胞的亲和力色谱法表达并纯化。使用微流动迁移率转移测定法对活性进行了验证。•使用底物磷酸化测定法来评估tern-701对人类组的选择性。使用CellTiter-Glo®评估了Tern-701抑制CML细胞系增殖的能力。使用固定摩尔组合和扩展的组合矩阵评估了Tern-701和主动位点TKI之间的协同作用,并使用CellTiter-Glo®测量细胞活力,并使用Bliss,组合指数(CI)和曲线移位分析进行了量化的相互作用。
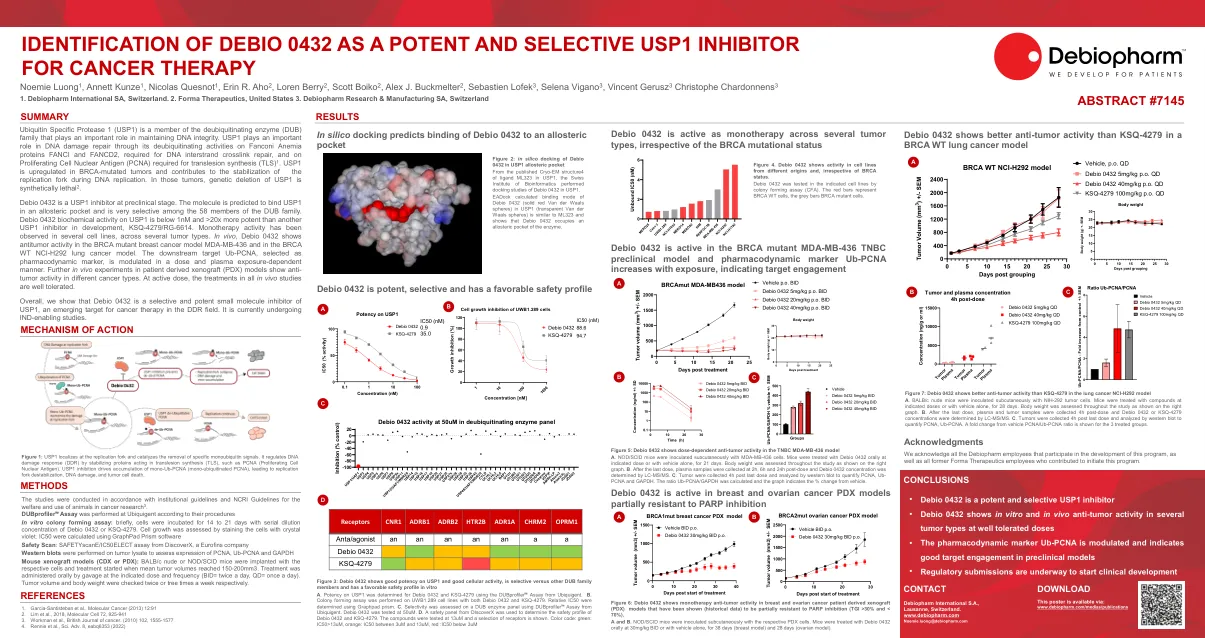
将Debio 0432识别为有效且有选择性的USP1 ...
Debio 0432是临床前阶段的USP1抑制剂。该分子预计将在变构袋中结合USP1,并且在DUB家族的58个成员中非常有选择性。DEBIO 0432 USP1上的生化活性低于1NM低于1NM,比其他USP1抑制剂(KSQ-4279/RG-6614)高20倍。单一疗法活性。在体内,Debio 0432显示了BRCA突变乳腺癌模型MDA-MB-436和BRCA WT NCI-H292肺癌模型中的抗肿瘤活性。以剂量和血浆暴露依赖性方式调节下游目标UB-PCNA,被选为药效标记。进一步的患者衍生异种移植物(PDX)模型的体内实验显示了不同癌症类型的抗肿瘤活性。在活性剂量时,所有体内研究中的治疗方法都得到很好的耐受性。
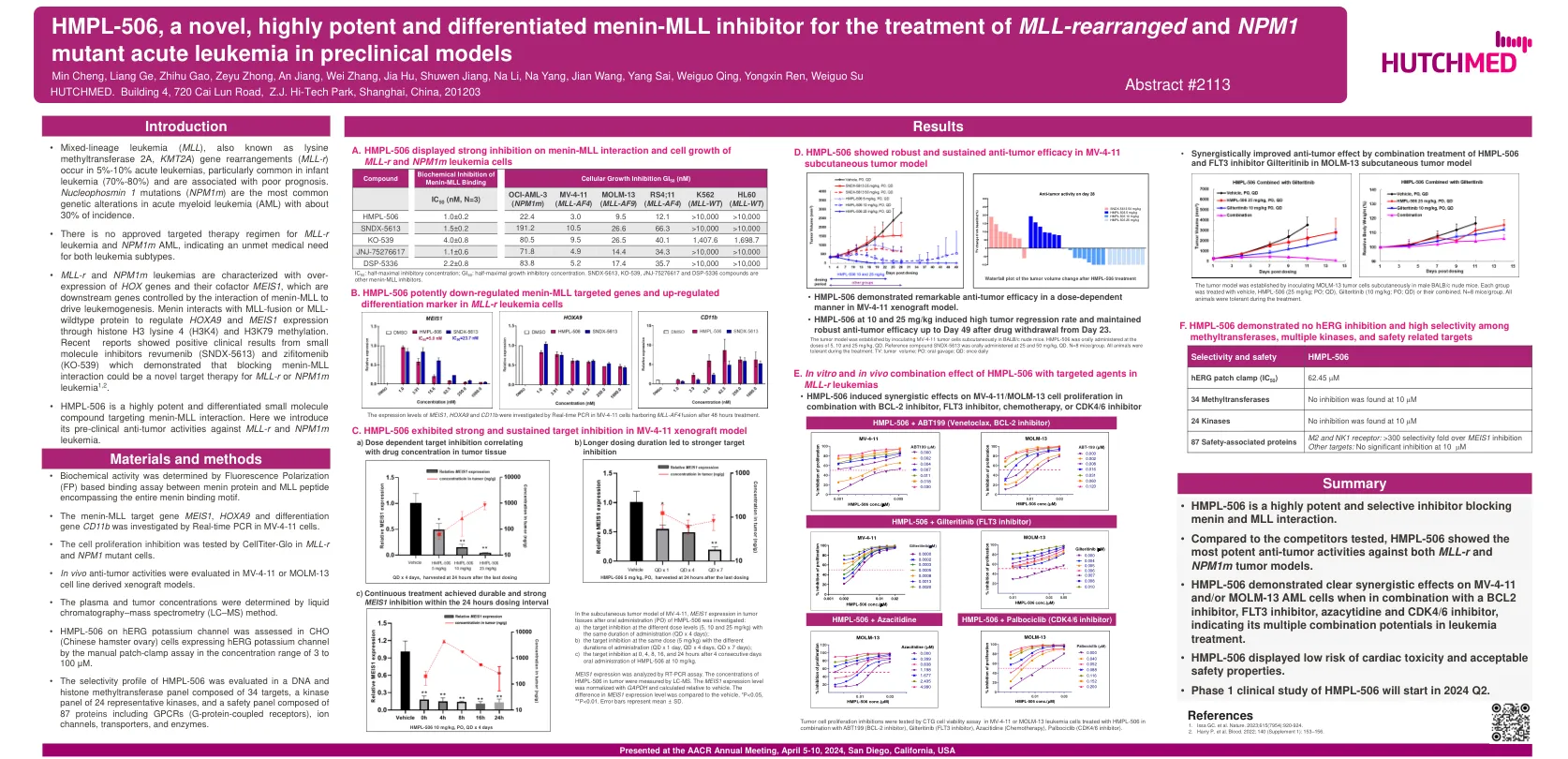
HMPL-506,一部小说,高度有效的和分化的Menin ...
HOX基因及其辅助因子Meis1的表达,它们是由Menin-Mll相互作用以驱动白血病发生的下游基因。Menin与MLL融合或MLL-野生型蛋白相互作用,通过组蛋白H3赖氨酸4(H3K4)和H3K79甲基化来调节HOXA9和MEIS1表达。最近的报道显示,小分子抑制剂rebumenib(SNDX-5613)和Zifitomenib(KO-539)的阳性临床结果,这些结果表明,阻断Menin-MLL相互作用可能是对MLL-R-R或NPM1M1M1M1M1M1M1M1M1M1,2的新型靶标治疗。

一种轻松的方法来产生...
本文档是公认的手稿版本的已发表作品,其最终形式以ACS药物化学信函出现,版权所有©美国化学学会后,在同行评审和发行者的技术编辑后。要访问最终编辑和发布的工作,请参见https://doi.org/10.1021/acsmedchemlett.0c00657

高效的活性药物成分可容纳
具有低效力材料的应用,其CPT(控制性能目标)<10µ/m³,即使以10kg质量转移量处理时,dusty也会产生25的过程得分。这适用于简单的环境压力柔性膜隔离器。现在,用更危险的API替换材料,其CPT <1.0µ/m³相同数量 - 相同的操作,过程得分上升至40。现在,应将控制装置升级到负压操作,H14 HEPA入口和排气以及压力损失警报系统。现在,用超高效力材料(如细胞毒素)代替API。体积传输很可能会减少,因为分配涉及少量。CPT <0.01µ/m³x<0.5kg转移大小 - 尘土飞扬的材料,过程得分上升至60。 div>要匹配60个过程得分,我们需要负压操作 + 2 X级H14排气过滤器 +压力损失警报,并且我们增加了每个开始周期上运行自动外壳泄漏测试(压力衰减测试)的能力。另外,我们反映了这种材料的高效力(0.01µ/m³cpt),我们设计了一个完全可容纳的隔离器。没有与HPAPI保持联系的不锈钢零件,并且可以在活动结束时使用所有HEPA过滤器“真空包装”的设计。

C-SRC激酶的选择性和有效的Protac降解器
C-SRC激酶Wuxiang Mao的选择性和有效的Protac Degrader,Nathalie M. Vandecan,Christopher R. Bingham,A Pui Ki Tsang,A Peter Ulintz,Brache ulintz,B Rachel Sexton,Daniel A. Bochar,Daniel A. Bochar,A Sofia D. Merajver,A Sofia D. Merajver,b and b and Matthew B.Softhew B. suellner*seellner*a。密歇根大学化学系,密歇根州安阿伯市930 N. University Ave.,48109。b。 密歇根大学内科系,1500 E. Medical Ave.,Ann Arbor,MI 48109。 使用链接到E3连接酶配体的dasatinib的摘要,我们确定了有效的双CSK/C-SRC Protac Degrader。 然后,我们用构象选择性类似物代替了dasatinib,稳定c-shelix c-Src的构象。 使用A c螺旋外配体,我们确定了一种对C-SRC有效且有选择性的Protac。 使用我们的C-SRC Protac,我们确定了与癌细胞增殖相比,C-SRC降解的药理优势。 引言蛋白激酶(PKS)在细胞信号传导和调节关键生物学过程(包括增殖,分化和凋亡)中起着至关重要的作用[1-3]。 对于许多激酶,对基因组敲低(例如siRNA)与激酶抑制剂的药理干预之间的细胞信号传导有不同的作用[4-6]。 基因组和药理学干预之间的这种断开是由于激酶的非催化功能仅被基因组敲低而破坏[4-6]。 因此,激酶指导的Protac代表了靶向激酶的潜在进步,该激酶非催化功能对于细胞信号很重要。密歇根大学化学系,密歇根州安阿伯市930 N. University Ave.,48109。b。密歇根大学内科系,1500 E. Medical Ave.,Ann Arbor,MI 48109。 使用链接到E3连接酶配体的dasatinib的摘要,我们确定了有效的双CSK/C-SRC Protac Degrader。 然后,我们用构象选择性类似物代替了dasatinib,稳定c-shelix c-Src的构象。 使用A c螺旋外配体,我们确定了一种对C-SRC有效且有选择性的Protac。 使用我们的C-SRC Protac,我们确定了与癌细胞增殖相比,C-SRC降解的药理优势。 引言蛋白激酶(PKS)在细胞信号传导和调节关键生物学过程(包括增殖,分化和凋亡)中起着至关重要的作用[1-3]。 对于许多激酶,对基因组敲低(例如siRNA)与激酶抑制剂的药理干预之间的细胞信号传导有不同的作用[4-6]。 基因组和药理学干预之间的这种断开是由于激酶的非催化功能仅被基因组敲低而破坏[4-6]。 因此,激酶指导的Protac代表了靶向激酶的潜在进步,该激酶非催化功能对于细胞信号很重要。密歇根大学内科系,1500 E. Medical Ave.,Ann Arbor,MI 48109。使用链接到E3连接酶配体的dasatinib的摘要,我们确定了有效的双CSK/C-SRC Protac Degrader。然后,我们用构象选择性类似物代替了dasatinib,稳定c-shelix c-Src的构象。使用A c螺旋外配体,我们确定了一种对C-SRC有效且有选择性的Protac。使用我们的C-SRC Protac,我们确定了与癌细胞增殖相比,C-SRC降解的药理优势。引言蛋白激酶(PKS)在细胞信号传导和调节关键生物学过程(包括增殖,分化和凋亡)中起着至关重要的作用[1-3]。对于许多激酶,对基因组敲低(例如siRNA)与激酶抑制剂的药理干预之间的细胞信号传导有不同的作用[4-6]。基因组和药理学干预之间的这种断开是由于激酶的非催化功能仅被基因组敲低而破坏[4-6]。激酶指导的Protac代表了靶向激酶的潜在进步,该激酶非催化功能对于细胞信号很重要。c-Src是一种酪氨酸激酶,是发现的第一个原始癌基因,并且在癌症中经常过表达[7-9]。虽然机制仍然鲜为人知[9],但C-SRC过表达的程度通常与恶性肿瘤的转移潜力相关,并且抑制C-SRC已被证明会降低小鼠的乳腺癌转移[10]。c-Src通过遗传敲低被验证为许多实体瘤的目标。然而,药理学抑制(无论是在临床还是临床前模型中)导致信号传导表型与遗传敲低不同[10]。敲低(例如,siRNA),三阴性乳腺癌(TNBC)和基底膀胱癌表现出降低和侵袭特性[10,11]。不幸的是,对C-SRC的小分子抑制剂的研究(包括:dasatinib,bosutinib和Ponatinib)未能概括从C-SRC的遗传敲低的强抗癌表型中,并且在诊所没有成功[10,11]。Protac提供了一种化学敲低的手段[12],因此我们有兴趣开发C-SRC的Protac。结果和讨论设计和评估C-SRC定向Protacs。为了识别C-SRC的PROTAC,我们设想将dasatinib(一种有效的C-SRC/ABL激酶抑制剂)与Thalidomide(Cereblon E3连接酶配体)结合在一起。据报道,基于dasatinib的daSatinib的protac是为了降解C-ABL和BCR-ABL,包括DAS-6-2-2-6(图1)[13]。我们希望DAS-6-2-2-6能够降解C-SRC,但是我们观察到Cal148细胞中C-SRC没有降解(18小时时100 nm)。与Protac文献一致[14],我们假设在DAS-6-2-2-6中发现的柔性且较长的接头不适合降解C-SRC。

在瞄准有效的植物固醇的计算机探索中
非小细胞肺癌Bishnu adhikari 1,Ashish Phuyal 2,Anuraj Phunyal 2,Nabin Upadhyaya 2,Amar Waiba 2,