XiaoMi-AI文件搜索系统
World File Search Systemrecombination

重新组合:使用同源重组的细菌基因工程
要进行重新组合,需要表达噬菌体重组系统的细菌菌株。噬菌体可以从其自己的启动子或异源调节启动子中表达。从其内源性噬菌体启动子中表达基因的基因赋予了紧密调控和坐标表达的优势,从而导致更高的重组频率。这是一个重要的优势,因为在许多情况下,高重组频率对于获得所需的重组至关重要。该单元的作者通常使用位于大肠杆菌染色体上的有缺陷的预言,最近将该预言的关键要素转移到了许多不同的质粒中(Thomason等,2005;另请参见评论)。在此预言系统中,噬菌体重组功能受到肉毒噬菌温度敏感的C I 857抑制剂的控制。在低温(30至34 C)下,重组基因会严重抑制,但是当细菌培养的温度转移到42 C时,它们会从P L启动子中高水平表达。在Datsenko和Wanner(2000)的质粒构建体中,重组基因位于质粒上,并从阿拉伯糖启动子表示。DATSENKO和WANNER质粒以及某些作者的质粒构建体具有DNA复制的温度敏感性。基于质粒的系统具有迁移率的优势 - 它们可以在不同的大肠杆菌菌株中转移到鼠伤寒沙门氏菌和其他革兰氏阴性细菌。但是,如果重新组合针对质量,则使用位于细菌染色体上的预言系统更容易。在诱导重组函数后,将修饰的DNA(DS)(DS)PCR产物或合成单链(SS)寡核苷酸(Oligo)引入到预防菌株中,通过电穿孔引入预防菌株中。通过选择或筛选存活电穿孔的细胞种群获得重新组件。一旦获得了所需的构建体,就可以通过另一个重组去除预言。或者,染色体上的工程等位基因可以通过P1转导将不同的宿主移动到另一个宿主中。具有温度敏感复制起源的质粒可能会因在适当温度下的生长而从重组菌株中丢失。

使用Effelsberg 100m望远镜寻找正电子重组线
这项工作提出了一种在电磁频谱的无线电范围内搜索重组线的方法,该方法通过几种单独的测量结果结合使用了灵敏度的显着提高。无线电区域中重组线的检测,尤其是正电子的重组线,是寻找暗物质的必不可少的灰烬,因为该线的强度揭示了观察到的位置的阳性量。这对于计算歼灭横截面的计算和有关暗物质的自我宣传的发现至关重要。这项工作中介绍的方法应用于13-15 GHz的频率范围,并使用2021年至2024年之间收集的数据,作为Telamon Project的一部分,Effelsberg的100 m射电望远镜。在NGC7027和W3OH校准源中检测到了这种方法的功能,并检测到氢重组线(H76α-H79α)。对于正电子(PS60α -PS62α)的重组线的振幅和流动,计算上限。在整个观察期的覆盖数据中,有一个3σ上限,河流密度为0。6-0。7 mjy,具体取决于重组线。此外,比较了NGC7027校准源的两种方法。单独确定NGC7027的每个重组线的上限的“单个”方法,提供3σ上限为5。1-7。2 mjy。6-4。0 mjy。相反,“组合”方法将三条线链接在一起,然后确定上限,3σ-上限为3。
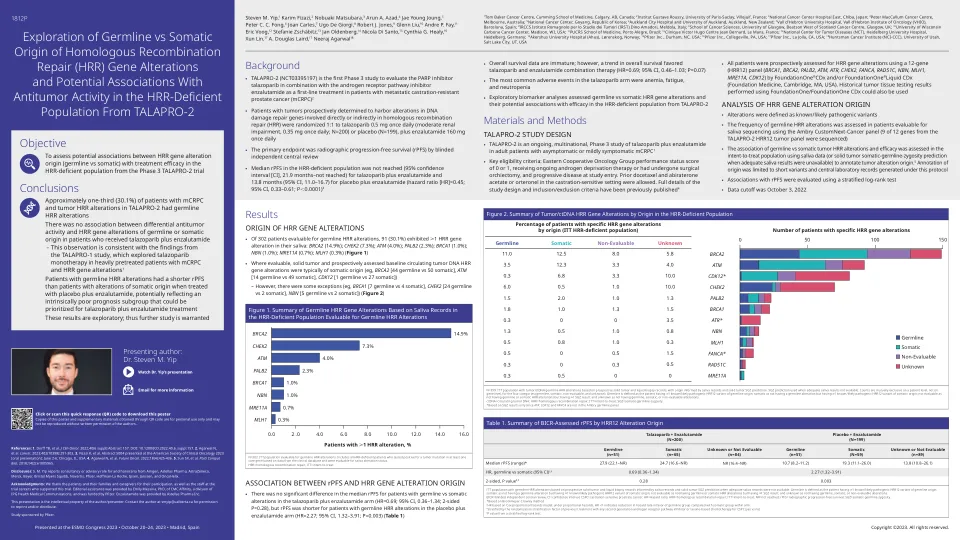
探索种系与同源重组的躯体起源
1汤姆·贝克癌症中心,卡明医学院,加拿大卡尔加里; 2法国维勒维夫大学,巴黎 - 萨克莱大学的Gustave Roussy研究所;日本千叶北部国家癌症中心医院3;澳大利亚墨尔本市彼得·麦卡伦(Peter MacCallum Cancer Center)4; 5号国家癌症中心,大韩民国戈阳; 6奥克兰市医院和奥克兰大学,新西兰奥克兰;西班牙巴塞罗那市Vall d'Hebron肿瘤学研究所(VHIO)Vall D'Hebron大学医院7; 8 Irccs iStituto romagnolo per lo Studio dei tumori(Irst)Dino Amadori,意大利梅尔多拉; 9英国格拉斯哥的苏格兰癌症中心的Beatson,格拉斯哥大学癌症科学学院9;威斯康星州麦迪逊市威斯康星州麦迪逊市威斯康星大学10号; 11 PUCRS医学院,巴西Porto Alegre; 12 Clinique Victor Hugo Center Jean Bernard,法国勒芒; 13德国海德堡海德堡大学医院国家肿瘤疾病中心(NCT);挪威洛伦斯科格(Lørenskog)14 Akershus大学医院(Ahus); 15美国北卡罗来纳州达勒姆市Pfizer Inc.; 16美国宾夕法尼亚州大学维尔市Pfizer Inc.; 17 Pfizer Inc.,美国加利福尼亚州拉霍亚; 18犹他大学,犹他州盐湖城,美国犹他州,亨斯曼癌症研究所(NCI-CCC)

表达异源重组酶的植物中同源重组的刺激
背景:CRISPR/Cas 和 TALEN 技术的进步激发了人们对植物基因编辑机会的兴奋。CRISPR/Cas 被广泛用于通过诱导靶向双链断裂 (DSB) 来敲除或修改基因,而双链断裂主要通过易出错的非同源末端连接或微同源介导的末端连接进行修复,从而导致可能改变或消除基因功能的突变。尽管此类突变是随机的,但它们发生的频率足以使有用的突变能够通过筛选定期识别。相比之下,用替代等位基因或具有特定特征修饰的拷贝替换整个基因的基因敲入目前还不常见。通过同源定向修复进行基因替换(或基因靶向)在高等植物中发生的频率极低,使得筛选有用事件变得不可行。通过抑制非同源末端连接和/或刺激同源重组 (HR) 可以增加同源定向修复。在这里,我们通过评估多种异源重组酶表达对烟草植物染色体内同源重组 (ICR) 的影响,为提高基因置换效率铺平了道路。结果:我们在含有高度敏感的 β -葡糖醛酸酶 (GUS) 型 ICR 底物的烟草转基因系中以不同的组合表达了几种细菌和人类重组酶。使用病毒 2A 翻译重编码系统实现了多种重组酶的协调同时表达。我们发现大多数重组酶在花粉中显著增加了 ICR,其中 HR 将由减数分裂期间发生的程序化 DSB 促进。DMC1 表达在初级转化体中产生了对 ICR 的最大刺激,其中一种植物的 ICR 频率增加了 1000 倍。对纯合 T2 植物系中的 ICR 的评估表明,ICR 增加了 2 倍到 380 倍,具体取决于表达的重组酶。相比之下,营养组织中的 ICR 仅适度增加,异源重组酶的组成性表达也降低了植物的育性。结论:异源重组酶的表达可以大大增加植物生殖组织中 HR 的频率。将此类重组酶表达与使用 CRISPR/Cas9 诱导 DSB 相结合可能是从根本上提高植物基因替换效率的途径。

从V(D)J重组到体细胞外显子改组
2016 年首次描述了通过插入编码额外结构域的大型序列获得特异性的抗体。在接触疟疾的个体中,来自白细胞相关免疫球蛋白样 1 ( LAIR1 ) 基因的外显子通过复制粘贴插入到免疫球蛋白重链编码区。几年后,发现了第二个例子,即来自白细胞免疫球蛋白样受体 B1 ( LILRB1 ) 基因的双外显子整合,该基因位于 LAIR1 附近。专门的高通量嵌合免疫球蛋白重链转录本表征揭示,来自遥远基因组区域(包括线粒体 DNA)的插入会导致人类抗体多样性。本综述描述了插入片段抗体的模式。在插入片段抗体生成背景下讨论了已知的 DNA 移动性方面的作用,例如基因组易位、基因转换和 DNA 脆弱性。最后,本综述介绍了为什么插入片段抗体在过去的库分析中被忽略,以及插入片段抗体如何有助于保护性免疫或自身反应性反应。

CRISPR/CAS通过同源重组的第3章基因组编辑
在整个细胞发育中,DNA可能遭受威胁基因组完整性和细胞存活的损害。最有害的病变之一是双链DNA断裂(DSB),因为它可能导致基因组信息的丢失。DSB可能自然发生在细胞代谢期间,也可能是由外部因素触发的(Deriano; Roth,2013)。无论哪种方式,这些损坏都会通过细胞立即修复,主要是通过两种途径:非同源末端连接(NHEJ)或同源指导修复(HDR)。与通过NHEJ进行修复不同,NHEJ仅将裂解的DNA的末端连接起来(请参阅第2章),HDR途径需要存在相同或非常相似的模板,即完整的序列,以准确地修复病变的DNA(Heyer等人,2010年)。提供用于HDR中使用的模板的可能性代表了通过同源重组(HR)途径进行基因编辑的关键元素,该途径可能被利用为几种新的繁殖技术(NBT)之一。

在酿酒酵母中交配类型切换过程中重组的中间体。
我们已经确定了从MATA到MATA的酵母交配型基因的同义转换的两个新型中间体。在HO核酸内切酶裂解后,观察到5'至3'的外核解消化,直到ho切割远端,产生了3'端的单链尾巴。在无法切换的RAD52应变中,此镜头更为广泛。令人惊讶的是,HO切割的近端受到保护,免受降解。这种稳定取决于无声复制供体序列的存在。通过定量应用聚合酶链反应(PCR)来鉴定第二个中间体。在MAT近端YA交界处出现之前,开关产物的YVA-MAT远端共价片段出现。未检测到MAT远端与HML远端序列的共价连接。我们建议,HO CUT远端的MAT DNA侵入完整的供体,并通过DNA合成扩展。在RAD52应变中阻止了此步骤。这些中间体与MAT开关的模型一致,在该模型中,HO切割的远端最初在链入侵和从供体中传递信息。关键词:重组机制/交配型/酵母/双链休息时间!rads2
将核酸外切酶与 Cas9 融合可增强毕赤酵母中的同源重组
HR 比 NHEJ 慢得多,NHEJ 可以从 DSB 事件中拯救更多细胞。NHEJ 几乎不需要或根本不需要末端切除来直接重新连接 DSB 末端。相比之下,HR 需要短距离切除和长距离切除 DSB 以及供体来实施修复过程。此外,其他蛋白质也可能是 HR 修复途径的限制因素 [18, 19]。我们在此发现,在同时删除两个基因和整合多个片段期间,将 MRE11 与 CAS9 融合可提高 CFU 数量

将质粒 DNA 和杆状病毒 DNA 共转染(同源重组)到 SF9 细胞中
1. 每瓶接种 5x10 6 个 SF9 细胞,另留一个瓶作为阴性对照。2. 按照 A 部分中概述的方法,使用以下体积和量:10uL 杆状病毒 DNA(0.1ug/uL)2ug 质粒 DNA 1mL 转染缓冲液 B 5mL 未添加的 Grace 培养基洗涤液 1mL 转染缓冲液 A 5mL 未添加的 Grace 培养基洗涤液 7mL TMN/FH

FEN1 内切酶作为同源重组缺陷人类癌症的治疗靶点
癌症治疗的合成致死策略利用癌症特异性基因缺陷来识别对肿瘤细胞存活至关重要的靶点。本文我们表明,RAD27/FEN1 编码瓣状内切酶 1 (FEN1),这是一种在 DNA 复制和修复中发挥作用的结构特异性核酸酶,与酿酒酵母基因组不稳定性基因具有最多的合成致死相互作用,是基于抑制剂的方法杀死同源重组 (HR) 缺陷癌症的可用药物靶点。研究表明,小分子 FEN1 抑制剂和 FEN1 小干扰 RNA (siRNA) 可选择性杀死 BRCA1 和 BRCA2 缺陷的人类细胞系,从而证实了 HR 缺陷癌症容易受到 FEN1 缺失的影响。此外,在小鼠中重现了对 FEN1 抑制的不同敏感性,小分子 FEN1 抑制剂降低了药物敏感但无耐药性癌细胞系中形成的肿瘤的生长。FEN1 抑制在敏感和耐药细胞系中均诱导了 DNA 损伤反应;然而,即使去除抑制剂,敏感细胞系也无法恢复和复制 DNA。尽管 FEN1 抑制在敏感细胞中将 caspase 激活到更高水平,但这种凋亡反应发生在 p53 缺陷细胞中,而泛 caspase 抑制剂无法阻断细胞杀伤。这些结果表明,FEN1 抑制剂具有治疗靶向 HR 缺陷癌症的潜力,例如由 BRCA1 和 BRCA2 突变和其他遗传缺陷引起的癌症。