XiaoMi-AI文件搜索系统
World File Search Systemreplica

深入研究人类数字孪生实现 - arXiv
摘要 — 人体数字孪生 (HDT) 是人体的数字复制品,可以反映完整的人体、人体的某些部分(如器官、流动、细胞)甚至人类行为。HDT 是从数字孪生 (DT) 制造概念推断出的人类特定复制应用,被定义为一种创建物理系统或流程的数字复制品的技术,旨在优化其性能并支持更准确的决策过程。本文的主要目标是通过浏览其基本概念、与 DT 的区别、现有发展和不同的应用领域,为读者提供 HDT 领域当前努力的全面概述。审查方法包括对科学文献、专利和工业计划的详尽审查,以及对正在进行和预见的 HDT 研究活动的讨论,强调其潜在的好处和局限性。关键词 — 数字孪生、人体数字孪生、建模

第四条 保护区住户指南
是,即使在“以旧换新”的情况下,也需要规划许可。如果住宅有原装窗户或门、优质的历史替代品或优质的现代复制品,拆除它们可能会被视为有害。鼓励修复,需要有合格从业人员出具证据,说明窗户或门无法合理修复,以支持拆除申请。如果可以接受替换,应使用与原始设计、尺寸、轮廓、细节、开启方式和材料完全相同的复制品。进行研究以帮助确定替换方案,或委托遗产专家协助可能会有所帮助。如果可能,应小心拆除和重新使用任何相关的玻璃。尽可能将传统家具转移到新窗户或门上。使用匹配的材料和饰面对窗户和门进行小规模修复。

用于在智能再制造中实现产品数字孪生的通用资产模型
摘要 数字孪生是传感组件、产品、流程、人员或系统的“实时”虚拟复制品。它通过嵌入式传感器和物联网从实体实时或近实时捕获信息,准确地复制被建模的实体。数字孪生在制造业中的许多应用已被研究。本文重点关注并致力于开发产品数字孪生,以减少再制造中数量、质量和需求不确定性的影响。本文从再制造特有的问题出发,推导出再制造产品数字孪生的功能要求,并提出了要再制造的通用资产的统一建模语言 (UML) 模型。该模型用于一个示例,强调需要将现有知识和数据转化为集成系统,以实现产品数字孪生,从而支持再制造流程规划。

数字孪生技术与高等教育
数字孪生是物理世界中物体、系统或环境的虚拟复制品,例如人类心脏、汽车或拥挤的大学校园。它通常可以通过传感器的实时数据进行更新,无论物理对应物如何变化,它都会随之变化。这让观察者可以即时详细地监控其变化。数字孪生技术还可以运行模拟,预测物体或系统在未来各个时间点的变化,例如心脏可能衰竭、汽车可能撞车或校园基础设施可能崩溃的时刻,并预测哪些干预措施可以改变其进程。“我们可以在虚拟世界中复制物理世界,并预测将会发生什么,”信息技术科学家迈克尔·格里夫斯 (Michael Grieves) 说,他在一年多前的一次会议上介绍了数字孪生概念。
![arxiv:2403.02477v1 [physics.optics] 4 2024年3月4日](/simg/4\4f4c8af1e582024a6822d83e72367198078dcf9a.webp)
arxiv:2403.02477v1 [physics.optics] 4 2024年3月4日
在光子环境中,复制对称性破坏(RSB)现象的理论建议[1,2]和实验演示[3,4]在复杂系统[5-9]领域产生了巨大影响。在巴黎的磁性自旋玻璃的方法中[10],当在相同条件下制备的系统的复制品可以在自由型的景观中与多个局部最小值一起到达不同的状态时,就会发生RSB。在使用随机激光器(RLS),光学模式和输入激发仪的振幅的磁性类比中,分别是旋转和内部温度的作用[1,2]。非常明显,正如Parisi [5]所说,RSB的实验证据是由Multimode RLS提供的,因为在这种情况下,可以观察模式的占用率,从而直接测量具有RSB签名的顺序参数函数。

细胞学 - 核 - 生物学 - IFRN
几个实验室已经能够将基本组成部分分离为酵母染色体的活性。这些成分包括丝粒,负责染色体运动的负责;一个这样称呼的自主复制序列,是导致细胞分裂和端粒之间染色体复制的重复,即完成染色体重复所需的染色体末端。当任何染色体的这些元素被分离和融合,然后重新引入酵母细胞时,它们在细胞分裂中的表现几乎像正常染色体一样。 ”

CSIR-IIIM在花卉文化任务下分发QPM
“节目让我想起了Bhimtal的访问。当人们朝naukuchiatal开车时,人们可以看到美丽的大玫瑰,尤其是攀岩玫瑰,这与NBRI花展期间所展示的相似。人们应该出来享受大自然。另一位访客Deepali教授说:“我喜欢由万寿菊和菊花制成的花艺和拉姆·曼迪尔的复制品。对于那些不能在1月22日拜访奉献仪式的人来说,这是一个完美的景点。”与NBRI和CIMAP的花朵一起,展览中还显示了47个参展商的收藏“ NBRI已经组织了60多年的演出,以在北部平原上流行玫瑰和丹花,以教育植物爱好者,” NBRI首席科学家S K Tiwari说。

公共云基础设施服务市场调查
纺织公司(“纺纱”)是根据大学研究生成的知识产权(IP)创建的初创公司。政府对英国有一个科学和技术超级大国的野心,大学与高科技纺纱公司之间的良好伙伴关系为经济增长和生产力做出了贡献。这篇评论研究了全球最成功的大学纺纱生态系统,以及英国境内,以确定支持衍生产品的最佳实践和机会,以支持更大的投资并在英国更快地增长。虽然评论的重点是科学技术,但我们注意到英国在人文,社会科学和艺术方面具有特殊的优势,这些优势已经显示出纺纱能力。这为英国提供了一个独特的机会,可以建立一个世界领先的创新生态系统,该生态系统从所有学术机构和学科中汲取灵感,并且比硅谷的复制品更雄心勃勃。任何世界一流的纺纱创作中心都需要以下正确的混合:

禁止和允许携带的物品 - AF.mil
• 武器(无论是否持有许可证),包括枪械、刀具、仿制武器或玩具武器(包括小刀和多功能工具,但须拆除)• 冷藏箱(例外:婴儿食品/药物/小尺寸 10x7x7 英寸)• 弹药或任何形式的爆炸物• 这是一次无烟航空展:包括电子烟/蒸汽笔• 产生火花的设备(例如:打火机、火柴等)• 宠物(协助残障人士的服务性动物除外)• 帐篷、便携式或临时遮阳篷• 大草坪椅(例外:小型折叠露营椅)• 自行车、滑板车、滑板、旱冰鞋• 狼牙棒、胡椒喷雾和个人电击枪/电击枪• 禁止携带外部酒精• 激光笔• 数字扫描仪• 玻璃容器• 非法药物/禁止携带管制物质• 烤架或烟花• 过度储存的物品,如公文包、行李袋和背包:必须小于 10x10x10(英寸)(例外:尿布袋、小钱包、小型相机包和腰包)• 无人机
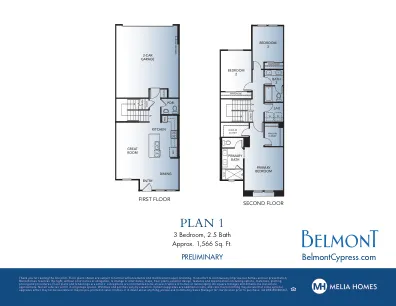
计划 1
感谢您阅读细则。显示的楼层平面图在最终确定时可能会进一步改进和修改。为了不断改进我们的房屋和展示,Melia Homes 保留更改或修改日期、地图、楼层平面图、外观设计、功能和规格(包括选项、材料、绘图、定价和程序)的权利,无需事先通知或承担义务。楼层平面图和效果图是艺术家的构想,并非旨在精确复制房屋或景观。所有平方英尺和尺寸(列出)均为近似值。并非所有车辆都适合所有车库空间。窗户和门廊因海拔高度而异。选项/升级需要额外费用,施工时间可能决定某些选项/升级可能不可用或可能在部分房屋中预先绘制。如果对任何事情有疑问,请在购买前询问社区销售经理以进行澄清。CA DRE #01881347。