XiaoMi-AI文件搜索系统
World File Search Systemserovar

通过单独对琼脂和其他培养基成分进行灭菌并降低琼脂浓度来改进倾注平板法
摘要 尽管倾注平板法在微生物质量控制中得到广泛应用,但它也存在某些缺点,包括必须在接种前融化培养基。在本研究中,通过使用较低浓度的琼脂(10 g/L)对培养基的制备进行了改进,琼脂在灭菌过程中与营养物质分离。在食品、化妆品和药品微生物质量控制中经常使用的培养基中评估了新方案,其中包括胰蛋白酶大豆琼脂 (TSA)、Sabouraud 4% 葡萄糖琼脂 (SDA) 和紫红胆汁葡萄糖琼脂 (VRBG)。与传统生产的培养基相比,改进后的培养基显著改善了 SDA 中酿酒酵母、金黄色葡萄球菌、肠道沙门氏菌亚种的生长。在 TSA 中可分离肠杆菌、鼠伤寒沙门氏菌和白色念珠菌,在 VRBG 中可分离大肠杆菌 ATCC 8739 和 ATCC 25922 以及鼠伤寒沙门氏菌。改良的 VRBG 对铜绿假单胞菌也更具选择性。至于物理化学性质,在 TSA 和 VRBG 中观察到 pH 值明显较低,在 TSA 中观察到强度值较低。将琼脂与培养基的其他成分分开灭菌,并将琼脂浓度降低至 10 g/L,可改善微生物生长,并提高倾注平板法中差异培养基的选择性。这些改进可以促进这种培养技术的自动化。

缺乏 RpoS 和二级信使 c-di-GMP 的 DIVA 疫苗株可用于预防猪沙门氏菌病
摘要 沙门氏菌病是欧盟第二大常见的食源性人畜共患病,猪是这种病原体的主要宿主。养猪生产中的沙门氏菌控制需要采取多种措施,其中可通过接种疫苗来减少流行血清型(如鼠伤寒沙门氏菌血清型)的亚临床携带和脱落。减毒活疫苗株在增强细胞介导免疫和允许通过口服途径接种方面具有优势。然而,这些疫苗的主要缺点是对异源血清型的交叉保护作用有限,并且会干扰感染的血清学监测。我们最近表明,减毒沙门氏菌菌株 (ΔXIII) 在鼠感染模型中对鼠伤寒沙门氏菌具有保护作用。ΔXIII 菌株含有 13 条染色体缺失,这使得它无法产生 sigma 因子 RpoS 和合成环二鸟苷酸 (c-di-GMP)。在本研究中,我们的目标是测试 ΔXIII 菌株对猪的保护作用,并研究使用 ΔXIII 是否可以区分已接种疫苗的猪和已感染的猪。结果表明,在断奶前仔猪口服 ΔXIII 疫苗可减少断奶和屠宰时的粪便脱落和回盲淋巴结定植,从而交叉保护仔猪免受鼠伤寒沙门氏菌的攻击。接种疫苗的猪在断奶时既没有粪便脱落,也没有疫苗菌株的组织持续存在,从而确保屠宰时不存在 ΔXIII 菌株。此外,ΔXIII 菌株中缺乏 SEN4316 蛋白,这使得开发血清学测试成为可能,从而区分感染动物和接种疫苗的动物 (DIVA)。

戊糖乳杆菌v390在酸性和胆汁条件下的活力及其抗菌活性和安全性评价
本研究使用 FYM27 和 R1492 引物进行 16S rRNA 基因分析,对 Lactiplantibacillus pentosus v390 进行分子鉴定。在 pH 2.5、3.5 和 4.5 的酸性条件下评估了菌株的生存力,并研究了对 0%、0.3%、0.5% 和 0.7% 浓度胆汁的抵抗力。评估了抗氧化活性、胆固醇吸收、疏水性、产生 DNase 酶的潜力、生物胺、溶血活性和对常见治疗性抗生素的耐药性。使用孔板和纸片扩散法检查了该菌株对致病菌(痢疾志贺氏菌、鼠伤寒沙门氏菌、大肠杆菌、金黄色葡萄球菌、单核细胞增生李斯特菌和枯草芽孢杆菌)的抗菌作用。结果表明,L. pentosus v390菌株在不同pH水平下均具有生存能力,但在pH 2.5下储存3小时后细菌数量下降。该菌株在不同胆汁盐浓度下均具有生长能力。L. pentosus v390对抗生素呋喃西林具有中等抵抗力,对萘啶酸和亚胺培南具有抗性,对万古霉素、庆大霉素、氯霉素、青霉素和环丙沙星等抗生素敏感。该菌株的疏水性、抗氧化活性(DPPH和ABTS)和胆固醇吸收率分别为46.50±0.38%、37.20±0.40%、39.90±0.45%和36.50±0.47%。未观察到该菌株产生DNase酶、生物胺或溶血活性。 L. pentosus v390 对革兰氏阳性菌表现出更强的抗菌作用。结果表明 L. pentosus v390 具有理想的益生菌特性,需要进一步研究以确认其在食品产品开发中的应用潜力。

使用外膜蛋白A,C和F
Salmonella Kentucky带来的全球公共卫生风险(S。肯塔基州)正在上升,特别是由于人类和动物种群中抗菌抗性基因的传播。这种血清在非洲普遍存在,已成为人类非脑性胃肠炎的显着原因。在这项研究中,我们使用了一种生物信息学方法来开发基于肽外膜蛋白A,C和F的基于肽的疫苗。肯塔基州。此外,我们采用了鼠伤寒沙门氏菌的鞭毛蛋白(FLIC)(s。鼠伤寒)作为增强疫苗有效性的佐剂。通过这种方法,我们确定了14个CD8+和7个CD4+ T细胞表位,这些表位预先限制为各种MHC I类和MHC II类等位基因。预计的表皮预计将在疫苗配方中使用时达到94.91%的覆盖率。此外,我们确定了七个高度免疫原性的线性B细胞表位和三个构象B细胞表位。然后,使用适当的接头将这些T细胞和B细胞表位连接起来,以创建多角色疫苗(MEV)。增强了肽构建体的免疫原性,从s。鼠伤寒在N末端包括在内。由此产生的MEV结构表现出高结构质量和有利的理化特性。通过Toll样受体1、2、4和5进行了分子对接研究,然后进行分子动力学模拟,表明疫苗受体综合在能量上是可行的,稳定的和健壮的。免疫模拟结果表明,MEV引起的显着反应,包括IgG,IgM,CD8+ T细胞,CD4+ T-细胞和各种细胞因子(IFN-γ,TGF-β,IL-2,IL-10和IL-12),以及抗原水平的显着降低。尽管有这些有希望的内部发现结果,但进一步验证

转基因生物:牛津疫苗组(20...
B2 与申请有关的转基因生物的一般描述 GMO 的描述 GMO 是野生型副伤寒沙门氏菌 (S. Paratyphi) A 9150 菌株的同源突变体。遗传改造的目的是构建一种带有 guaBA 操纵子和 clpX 基因缺失的改良 S. Paratyphi A 9150 菌株,以产生生长缺陷的减毒 S. Paratyphi A 菌株 (CVD 1902)。GMO (CVD 1902) 将用于研究其作为减毒活口服疫苗在受控人类感染模型中预防肠热病的价值。CVD 1902 副伤寒沙门氏菌 A 血清型活口服疫苗是由野生型亲本菌株 S. Paratyphi A 9150 构建的。使用改良的 Lambda Red 介导的定点诱变程序进行缺失。删除了两个基因序列:guaBA 染色体操纵子(编码远端从头鸟嘌呤核苷酸生物合成途径中使用的两种酶)和 clpX 基因(编码分子伴侣 ATPase,与 clpP 编码的丝氨酸蛋白酶一起发挥作用,形成参与各种代谢过程的复合物)。clpX 缺失突变的表型后果之一是鞭毛的过度表达,这也可能增强 GMO 菌株的免疫原性,因此有助于其作为减毒活疫苗的适用性。从研究中获得的信息将用于指导疫苗设计和开发,从而可能影响公共卫生干预策略。应用描述在这项临床研究中,我们建议调查 GMO CVD 1902 作为减毒活疫苗在 S. Paratyphi A 人类攻击模型中预防副伤寒感染的有效性。使用野生型 S 的 S. Paratyphi A 感染的人感染模型。副伤寒甲型(NVGH308 株)已在牛津疫苗组(英国牛津大学)建立。牛津疫苗组(英国牛津大学)一直在进行
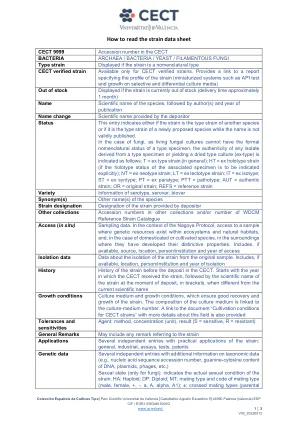
如何读取应变数据表
CECT 9999 CECT细菌中的登录数 /细菌 /细菌 /酵母 /丝状真菌型应变,如果应变是命名型型CECT CECT CECT验证的菌株,仅可用于CECT经过验证的菌株。提供了指定应变概况(小型化系统(如API测试和选择性和差异培养媒体上的增长)的概况的报告链接,如果库存出现的库存显示,如果劳力目前缺货(大约1个月),该物种的名称是该物种的科学名称,则应通过作者名称和本性名称的定期来指示。物种虽然该名称未有效出版。在真菌的情况下,由于活真菌培养物不能具有类型标本的形式命名命名状态,因此从类型标本中得出的任何分离物的真实性或产生干燥类型培养物(EX-Type)的真实性如下:T = t = ex type株(通常); HT = Ex Holotype菌株(如果要明确指示相关样品的整型状态); nt = ex neotype菌株; lt =外型应变; it = ex iSotype; st = ex syntype; pt = ex Paratype; ptt = pathotype; aut =正宗应变;或=原始应变;参考=参考应变品种,血清型,血清,血清,Biovar同义词的其他名称的其他名称名称是由存款人提供的应变的菌株名称名称,其他集合中的其他集合登录号和/或WDCM参考菌株分类目录访问(原位)采样数据。培养基的组成与培养物中的数量有关。在名古屋方案的背景下,在生态系统和自然栖息地中存在遗传资源的样本,以及在驯养或耕种物种的情况下,在他们开发出独特特性的环境中。包括(如果有),包括来源,位置,人员/机构和访问年份隔离数据数据,涉及与原始样品隔离的隔离。包括(如果有),包括位置,人员/机构和隔离的年度历史历史记录在cect中。从CECT收到压力的年份开始,然后在存款时,在括号中的菌株的科学名称,当时与当前的科学名称生长条件培养培养基和生长条件不同,这确保了应变的良好恢复和生长。还提供了有关该领域的更多详细信息的文档“培养条件”的链接

Accugen检测常见的临床致病微生物
Acinetobacter Baumannii, Staphylococcus capnocytophaga Haemolytica, Pseudomonas fluorescens, Staphylococcus horses, Actinomyces Israelii, Staphylococcus Epidermidis, Capnocytophaga Ochracea, Pseudomonas Mosselii, Streptobacillus moniliformis, Bordetella tunnels,葡萄球菌血液溶血,囊孢子虫,pseudomonas putida,链球菌,Gallolyticus,Burkholderia cepacia,葡萄球菌,弯曲球菌,弯曲球菌Ococcus沙门氏菌肠道SSP。 div>Acinetobacter Baumannii, Staphylococcus capnocytophaga Haemolytica, Pseudomonas fluorescens, Staphylococcus horses, Actinomyces Israelii, Staphylococcus Epidermidis, Capnocytophaga Ochracea, Pseudomonas Mosselii, Streptobacillus moniliformis, Bordetella tunnels,葡萄球菌血液溶血,囊孢子虫,pseudomonas putida,链球菌,Gallolyticus,Burkholderia cepacia,葡萄球菌,弯曲球菌,弯曲球菌Ococcus沙门氏菌肠道SSP。 div>

沙门氏菌Shigella琼脂/XLD琼脂
1。预期的用途检测和分离革兰氏阴性肠病原体,尤其是人类临床标本和其他标本中的志贺氏菌和沙门氏菌。革兰氏阴性肠病原体(尤其是志贺氏菌和沙门氏菌)的Shalmella shigella琼脂/XLD琼脂。沙门氏菌琼脂/XLD琼脂的功能是支持症状患者的诊断,表明革兰氏阴性肠病原体,尤其是Shigella属和沙门氏菌的病原体潜在感染。沙门氏菌是食物中毒的一些最常见的病因。这些微生物的致病性从一种血清变化到另一种血清,并且在同一亚种中可能会有所不同。一些血清造成了侵入性疾病,但也有一些造成自限性食物中毒的血清疾病。沙门氏菌肠subsp的最孤立的血清。肠道是S. enteritidis,S。Typhimurium,S。Virchow,S。Hadar或S. iftantis。Shigella属包括四种:S。dysenteriae,s。Flexneri,S。Boydii和S. Sonnei。所有物种都是强制性的病原体,并引起细菌痢疾。2。手术沙门氏菌琼脂的原理胆汁盐,孔雀石绿色和柠檬酸钠的存在抑制了除沙门氏菌和志贺氏菌以外的革兰氏阳性微生物和肠杆菌的生长。由于添加乳糖,肠杆菌的分化是可能的。乳糖发酵细菌会产生酸并形成红色菌落,这是由于中性红色的pH指示剂。相反,乳糖非发酵微生物形成无色菌落。柠檬酸铁是硫化氢产生的指标。沙门氏菌产生硫代硫酸盐还原酶,该酶释放出存在于硫代硫酸钠中的硫化物分子。这些分子与氢离子结合,形成H 2 S,与柠檬酸铵反应。这种反应导致形成沉淀物,可见在细菌菌落中心的黑点。XLD琼脂酵母提取物是培养基中养分的来源。脱氧胆酸钠的存在抑制了革兰氏阳性细菌的生长。由于三个指示系统,细菌的分化是可能的: - 乳糖,木糖和蔗糖与苯酚红(这是pH指示剂) - - 盐酸l-赖氨酸盐和苯酚红色, - 硫代硫酸钠和柠檬酸铁硫酸盐。木糖的发酵降低了培养基的pH值,并使其从红色变为黄色。包括沙门氏菌在内的大多数肠道病原体能够发酵木糖,从而导致培养基的酸化。由于志贺氏菌的细菌是乳糖的非发酵,因此不会产生酸,因此会形成红色菌落。赖氨酸允许将沙门氏菌细菌与其他非致病细菌区分开。一旦木糖耗尽,沙门氏菌细菌在脱羧过程中利用L-赖氨酸,这将培养基的pH水平改变为碱。为防止赖氨酸阳性大肠菌群,乳糖和蔗糖的类似pH水平的类似回归,以产生多余的酸。氯化钠保持渗透平衡。柠檬酸铵是硫化氢生产的指标。沙门氏菌产生硫代硫酸盐还原酶,该酶释放出存在于硫代硫酸钠中的硫化物分子。这些分子与氢离子结合形成H 2 s,与柠檬酸铁反应形成沉淀物,可见在细菌菌落中的黑色中心。产生H 2 S的非致病细菌不脱羧L-赖氨酸。因此,它们产生的酸反应阻止了菌落的变化。