XiaoMi-AI文件搜索系统
World File Search SystemsgRNA

从可解释的人工智能中了解 CRISPR-Cas9 sgRNA 效率的量子生物学见解
1 计算和预测生物学,生物科学,橡树岭国家实验室,美国田纳西州橡树岭 2 田纳西大学诺克斯维尔分校布雷迪森跨学科研究与研究生教育中心,美国田纳西州橡树岭 3 合成生物学,橡树岭国家实验室,美国田纳西州橡树岭 4 计算科学与工程,橡树岭国家实验室,美国田纳西州橡树岭 本稿件由 UT-Battelle, LLC 根据与美国能源部签订的合同编号 DE-AC05- 00OR22725 撰写。美国政府保留;并且出版商在接受文章发表时,承认美国政府保留非独占的、已付费的、不可撤销的全球许可,可以为美国政府的目的出版或复制本稿件的已出版形式,或允许他人这样做。能源部将根据能源部公共访问计划 ( http://energy.gov/downloads/doe-public-access-plan ) 向公众开放这些联邦资助研究的成果。摘要:CRISPR-Cas9 工具已经彻底改变了实验室的基因操作能力。经验法则仅针对少数模型生物建立,而 sgRNA 效率的机制基础仍然知之甚少。这项工作建立了一个使用量子化学张量生成的新特征集和新公共资源,用于解释和预测 sgRNA 效率。sgRNA 效率的特征工程是使用可解释的人工智能模型;迭代随机森林 (iRF) 执行的。通过对大肠杆菌 sgRNA 的位置特异性序列的定量属性进行编码,我们确定了细菌物种中 sgRNA 设计的重要性状。此外,我们还表明,将位置编码扩展到碱基对、二聚体、三聚体和四聚体序列的量子描述符可以捕获目标 DNA 局部和邻近核苷酸中复杂的相互作用。这些特征凸显了大肠杆菌和智人基因组之间 CRISPR-Cas9 sgRNA 动力学的差异。这些新颖的 sgRNA 编码极大地增强了我们对 CRISPR-Cas9 机制中涉及的复杂量子生物过程的理解。

指南 - IT SGRNA体外转录套件产品组件列表
通知购买者我们的产品仅用于研究目的。它们不得用于任何其他目的,包括但不限于用于药物,体外诊断目的,治疗剂或人类的使用。我们的产品不得转让给第三方,转售,转售或用于制造商业产品或未经Takara Bio USA,Inc。的书面批准,为第三方提供服务。

非洲绿猴(African Green Monkey)全基因组CRISPR sgRNA ...
目的基因 sgRNA 数目: 64853 ;阴性对照 sgRNA 数目: 2000 ; sgRNA 大小: 20bp

用于人类基因中 CRISPRa sgRNA 设计的网络工具的计算机性能分析
血管生成基因过表达已成为众多血管再生基因治疗项目的主要策略。然而,大多数项目在临床试验中都失败了。CRISPRa 技术以最高的效率和安全性在 sgRNA 的识别基础上提高基因过表达水平。CRISPick 和 CHOP CHOP 是用于预测 sgRNA 的最广泛使用的 Web 工具。我们的研究目的是分析这两个平台对于涉及不同人类参考基因组(GRCH 37 和 GRCH 38)的血管生成基因(VEGFA、KDR、EPO、HIF-1A、HGF、FGF、PGF、FGF1)的 sgRNA 设计的性能。从不同方面分析了这两个工具提出的排名前 20 的 sgRNA。与 sgRNA 结合位点相关的 DNA 曲率没有发现显著差异,但使用 CRISPick 时,sgRNA 预测的靶向效率显著更高。此外,同一平台在 EPO、EGF、HIF-1A、PGF 和 HGF 中的平均排名变化较大,而在 KDR、FGF-1 和 VEGFA 中未达到统计显著性。不同平台之间排名位置的重排分析也不同。CRISPick 被证明在与更完整的基因组相关的最佳 sgRNA 建立方面更准确,而 CHOP CHOP 显示出更窄的分类重排。2022 由 Elsevier BV 代表计算和结构生物技术研究网络出版。这是一篇根据 CC BY-NC-ND 许可协议 ( http://creativecommons.org/licenses/by-nc-nd/4.0/ ) 开放获取的文章。

sgRNA 的位点特异性和酶促交联可实现波长可选的光激活控制 CRISPR 基因编辑
化学交联能够快速识别 RNA-蛋白质和 RNA-核酸分子间和分子内相互作用。然而,目前尚无方法能够位点特异性和共价交联 RNA 内两个用户定义的位点。在这里,我们开发了 RNA-CLAMP,它能够位点特异性和酶促交联(夹紧)RNA 内两个选定的鸟嘌呤残基。分子内夹紧会破坏正常的 RNA 功能,而随后对交联剂进行光裂解会恢复活性。我们使用 RNA-CLAMP 通过光裂解交联剂夹紧 CRISPR-Cas9 基因编辑系统的单向导 RNA (sgRNA) 内的两个茎环,完全抑制编辑。可见光照射会裂解交联剂并以高时空分辨率恢复基因编辑。设计两种对不同波长的光有响应的光裂解接头,可以在哺乳动物细胞中实现基因编辑的多路复用光激活。这种光激活的 CRISPR-Cas9 基因编辑平台受益于无法检测的背景活动,提供激活波长的选择,并具有多路复用功能。

考虑 tracrRNA 序列中的细微变化可改善 CRISPR 筛选的 sgRNA 活性预测
CRISPR 技术是研究基因组功能的强大工具。为了帮助从众多可能的选项中挑选出对目标靶标具有最大功效的 sgRNA,几个研究小组开发了预测 sgRNA 靶向活性的模型。尽管多种 tracrRNA 变体通常用于筛选,但现有的模型在提名 sgRNA 时都没有考虑到这一特征。在这里,我们开发了一个靶向模型,规则集 3,它可以对多种 tracrRNA 变体做出最佳预测。我们在一个新的 sgRNA 数据集上验证了规则集 3,该数据集涵盖了必需和非必需基因,与之前的预测模型相比有显著的改进。通过分析 tracrRNA 变体之间 sgRNA 活性的差异,我们表明 Pol III 转录终止是 sgRNA 活性的重要决定因素。我们期望这些结果能够提高 CRISPR 筛选的性能,并为未来对 tracrRNA 工程和 sgRNA 建模的研究提供参考。
在CRISPR/CAS9应用的单个分析中,SGRNA和Cas9 mRNA的高分辨率表征
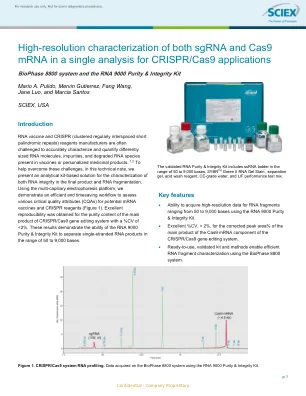
RNA疫苗和CRISPR(簇簇的定期间隔短的短粒子重复重复序列)制造商通常会挑战制造商,以准确表征和量化不同尺寸的RNA分子,杂质和降解的RNA物种,以及疫苗或个性化药物产品中的降解RNA物种。1,2为了帮助克服这些挑战,在本技术说明中,我们提出了一种基于分析套件的解决方案,用于表征最终产品中的RNA完整性和RNA片段化。使用多毛细管电泳平台,我们展示了有效且延时的工作流程,以评估潜在的mRNA疫苗和CRISPR试剂的各种关键质量属性(CQA)(图1)。对于CRISPR/CAS9基因编辑系统的主要产物的纯度含量获得了出色的可重复性,CV <2%。这些结果证明了RNA 9000纯度和完整性套件在50至9,000个碱基范围内将单链RNA产物分离的能力。

使用全基因组 CRISPRi 筛选来探测遗传修饰因子的双 sgRNA 文库设计
© 作者 2023。开放存取 本文根据知识共享署名 4.0 国际许可进行授权,允许以任何媒体或格式使用、共享、改编、分发和复制,只要您给予原作者和来源适当的信任,提供知识共享许可的链接,并指明是否做了更改。 本文中的图片或其他第三方资料包含在文章的知识共享许可中,除非资料的致谢中另有说明。 如果资料未包含在文章的知识共享许可中,且您的预期用途不被法定规定允许或超出允许用途,则需要直接从版权所有者处获得许可。 要查看此许可证的副本,请访问 http://creativecommons.org/licenses/by/4.0/。知识共享公共领域贡献豁免(http://creativecommons.org/publicdomain/zero/1.0/)适用于本文中提供的数据,除非数据来源中另有说明。
有效地递送活性Cas9蛋白和靶标特异性SGRNA到广泛的细胞类型
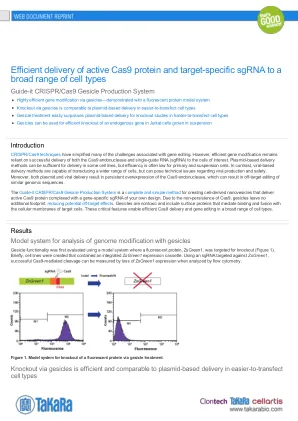
最后,在悬浮液中生长的Jurkat细胞中,评估了吉赛克的敲除生理相关的内源基因。敲除靶向的CD81,该CD81编码在许多哺乳动物细胞中表达的细胞表面蛋白,并与丙型肝炎,HIV和流感发病机理有关。jurkat细胞用Cas9和CD81特异性SGRNA的表达质粒共转染,或用靶向CD81的Cas9 -Sgrna RNP复合物预加载的gesiles处理。通过抗体标记评估CD81的基因敲除效率,然后进行流式细胞仪分析(图4)。基于质粒的递送导致效率非常低。只有7%的细胞损失CD81表达。相反,吉塞拉(Gesicles)具有很高的效率,其中76%的Jurkat细胞缺乏可检测到的CD81水平。因此,吉质的表现优于基于质粒的技术,具有在难以转化细胞中有效靶向内源基因的能力。

在微生物 CRISPR 干扰中使用缩短和随机 sgRNA 文库筛选新靶基因
摘要:CRISPR 干扰(CRISPRi)筛选已用于使用单分子向导 RNA(sgRNA)文库识别与特定表型相关的靶基因。在 CRISPRi 筛选中,包含原始靶标识别序列的随机 sgRNA 文库的大小很大(∼ 10 12 )。在本文中,我们证明 sgRNA 中的靶标识别序列(TRS)的长度可以从原来的 20 个核苷酸(N 20 )缩短到 9 个核苷酸(N 9 ),这仍然足以使 dCas9 抑制大肠杆菌木糖操纵子中的靶基因,无论其与启动子还是开放阅读框区结合。基于结果,我们构建了 TRS 长度 5′ 缩短的随机 sgRNA 质粒文库,并通过对从 Xyl − 表型细胞中纯化的 sgRNA 质粒进行桑格测序来识别木糖代谢靶基因。接下来,利用随机 sgRNA 文库筛选靶基因,以增强合成大肠杆菌细胞中紫色素的产生。通过分析深紫色菌落中 sgRNA 质粒中的 TRS 冗余度,选择了 17 个靶基因。其中,已知有 7 个基因(tyrR、pykF、cra、ptsG、pykA、sdaA 和 tnaA)可增加细胞内 L-色氨酸池(紫色素的前体)。17 个细胞中每个靶基因有一个缺失,紫色素的产量显著增加。这些结果表明,使用缩短的随机 TRS 文库进行 CRISPRi 可以简单且经济高效地进行基于表型的靶基因筛选。关键词:CRISPR 干扰、失活 Cas9、随机文库、缩短的 sgRNA、靶标识别序列、紫色素、基于表型的靶标筛选■简介