XiaoMi-AI文件搜索系统
World File Search SystemsgRNA

确认请求 21-356-01cr
图 l 中所示的 sgRNA tl 是根据几个 CB|-Deleted 算法设计和选择的,以最大限度地提高靶标特异性并降低二次靶标编辑的可能性。Young 等人 (20L9) 证明,通过生物信息学预测为特异性的 sgRNA 不会导致玉米中的二次靶标突变。用于创建减少褐变香蕉的两个选定 sgRNA 被用作 Cas-OFFinder 工具 (http://www.rgenome,net/cas-offinder/) (Bae S. 等人,2014) 的输入,该工具提供了潜在的

汇集表型 CRISPR 筛选能够识别用于癌症治疗的协同药物组合。
图 1:CRISPR 死亡筛选设置。在死亡细胞群中确定 sgRNA 丰度,从而可以识别在泛 PI3K 抑制剂 GDC-0941 存在下诱导细胞死亡的 sgRNA,并可以识别适合联合治疗的靶标。

使用 CRISPR-Cas9 核糖核蛋白复合物编辑康乃馨中的乙烯生物合成基因。
摘要 本研究利用CRISPR/Cas9核糖核蛋白(RNP)复合体系统对康乃馨乙烯(ET)生物合成基因[1-氨基环丙烷-1-羧酸(ACC)合成酶1(ACS1)和ACC氧化酶1(ACO1)]进行编辑。首先,验证靶基因(ACS1和ACO1)的保守区域,以生成不同的单向导RNA(sgRNA),然后使用体外切割试验验证sgRNA特异性切割靶基因的能力。体外切割试验表明,sgRNA在切割各自的靶区域方面具有很高的效率。将sgRNA:Cas9复合物直接递送到康乃馨原生质体中,并对原生质体中的靶基因进行深度测序。结果表明,sgRNA 适用于编辑 ET 生物合成基因,因为 ACO1 的突变频率范围为 8.8% 至 10.8%,ACS1 的突变频率范围为 0.2–58.5%。在对用 sgRNA:Cas9 转化的原生质体产生的愈伤组织中的目标基因进行测序时,在 ACO1 中发现了不同的 indel 模式(+ 1、- 1 和 - 8 bp),在 ACS1 中发现了不同的 indel 模式(- 1、+ 1 和 + 11)。这项研究强调了 CRISPR/Cas9 RNP 复合物系统在促进康乃馨 ET 生物合成的精确基因编辑方面的潜在应用。关键词 愈伤组织,CRISPR/Cas9,乙烯生物合成基因,Indel 模式,体外裂解,原生质体

农杆菌和单个 Cas9-sgRNA 转录系统介导的多年生黑麦草高效基因编辑
基因组编辑技术为多年生黑麦草(一种全球重要的牧草和草坪草种)的遗传改良提供了强有力的工具。关于多年生黑麦草基因编辑的唯一出版物使用基因枪进行植物转化,并使用基于双启动子的 CRISPR/Cas9 系统进行编辑。然而,它们的编辑效率很低(5.9% 或只产生了一株基因编辑植物)。为了测试玉米泛素 1 (ZmUbi1) 启动子在多年生黑麦草基因编辑中的适用性,我们制作了 ZmUbi1 启动子:RUBY 转基因植物。我们观察到 ZmUbi1 启动子在芽再生之前的愈伤组织中活跃,这表明该启动子适用于多年生黑麦草中的 Cas9 和 sgRNA 表达,以高效生产双等位基因突变植物。然后,我们使用 ZmUbi1 启动子来控制多年生黑麦草中的 Cas9 和 sgRNA 表达。Cas9 和 sgRNA 序列之间的核酶切割靶位点允许在转录后产生功能性 Cas9 mRNA 和 sgRNA。使用农杆菌进行遗传转化,我们观察到在多年生黑麦草中编辑 PHYTOENE DESATURASE 基因的效率为 29%。DNA 测序分析表明,大多数 pds 植物含有双等位基因突变。这些结果表明,由 ZmUbi1 启动子控制的单个 Cas9 和 sgRNA 转录单元的表达为产生多年生黑麦草的双等位基因突变体提供了一种高效的系统,并且也适用于其他相关草种。

CRISPR-Cas9 筛选用于目标识别。
图 2:PLX-4032 治疗后 sgRNA 计数分布的变化。log2 转换的 sgRNA 计数的箱线图和须线图(基线 - 抗生素选择后的转导细胞;DMSO 和 PLX-4032 - 筛选终点的细胞(治疗 14 天))。箱线图的范围从第一四分位数到第三四分位数,并以黑线显示中位数。此外,下部和上部相邻值显示为须线,异常值显示为圆圈。

CRISPR使用霓虹灯电穿孔对永生的细胞系编辑
该方案描述了如何将核糖核蛋白(RNP)复合物组成,这些复合物由纯化的CAS9核酸酶复制,用化学改良的合成单导剂RNA(SGRNA)复制到永生的粘附或悬浮液中。包括一个敲门选项。RNP递送是使用Thermo Fisher Neon™转染系统完成的。包括各种细胞类型的电穿孔设置的参考。化学修饰的SGRNA旨在抵抗可导致细胞死亡的外核酸和先天的细胞内免疫级联反应。Editco化学修改的合成SGRNA具有特殊的纯度,并且始终驱动高编辑频率。
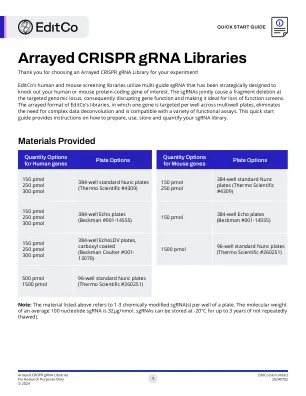
Arryed Crispr grna库
感谢您为您的实验选择Arrayed Crispr Grna库!Editco的人类和小鼠筛选库利用多指导SGRNA的策略性设计来消除您感兴趣的人类或小鼠蛋白质编码基因。SGRNA在靶向基因组基因座处共同引起片段缺失,因此破坏了基因功能并使其非常适合丧失功能筛选。Editco库的阵列格式,其中一个基因在多井板上的每个孔都被靶向,从而消除了对复杂数据反卷积的需求,并且与各种功能测定兼容。此快速启动指南提供了有关如何准备,使用,存储和量化SGRNA库的说明。

NDP 外显子 2 区域 KO 报告模板...
图 S01:CRISPR/Cas9 编辑人类 iPSC。(A) 使用双 sgRNA(sg1 和 sg2)靶向外显子 2 两侧的区域来生成 HDR 介导的 NDP KO-GFP 报告系,使用带有 KO 报告模板的 PUC57 载体,由外显子 2 (5'UTR)-eGFP-P2A 盒组成,两侧是同源臂 (HA),其中包括 PAM 位点突变和用于质粒线性化的 sgRNA 切割位点 (sg1-c 和 sg2-c)。在此过程中还生成了 NHEJ 介导的 NDP KO (NDP Δ exon2) 以及 NDP WT 同源克隆。(B) 使用类似方法生成 NDP WT-GFP 报告系。外显子 3 的 CDS 两侧的双 sgRNA(sg3 和 sg4)和含有修复模板的质粒,由外显子 3(CDS)-V5-P2A- eGFP 盒组成,两侧是同源臂,包括 PAM 位点突变和用于质粒线性化的 sgRNA 切割位点(sg3-c 和 sg4-c)。sgRNA,小向导 RNA;HDR,同源定向修复;KO-GFP,敲除报告基因;KO,敲除;WT,野生型;PAM,protoscpacer 相邻基序;UTR,非翻译区;CDS,编码序列;HA,同源臂;eGFP,增强型绿色荧光蛋白;P2A,猪 teschovirus-1 2A 自切割肽;V5,V5 标签。

新型 CRISPR 工具在心血管研究和医学中的应用
基因组编辑技术的发展彻底改变了生物医学研究,特别是自从 CRISPR/Cas9(成簇的规律间隔的短回文重复序列/CRISPR 相关蛋白 9)技术引入以来。该系统最初是在细菌免疫反应中发现的 1,随后应用于真核生物基因组编辑 2 – 4。CRISPR/Cas9 系统的简单性使基因组编辑比传统的 DNA 编辑技术更容易获得和更容易。作为一种基因组编辑工具,该系统由两个基本成分组成:一种切割 DNA 链的内切酶 Cas9,以及一种单向导 RNA(sgRNA),其中包含用于识别目标 DNA 区域的特定序列。值得注意的是,细菌中的内源性 Cas9 系统有两种 RNA 成分(CRISPR RNA(crRNA)和反式激活 crRNA); CRISPR/Cas9 工具中的 sgRNA 是由 crRNA 和反式激活 crRNA 人工改造而成。当 Cas9 和 sgRNA 被递送到细胞中时,产生的 Cas9-sgRNA 复合物被导向目标基因组位点,在那里产生 DNA 中的双链断裂 (DSB)。然后通过内源性 DNA 修复机制修复 DSB,从而实现基因敲除或敲入 5 。

利用-QM-MM和分子型模拟 -
图2。DNA,SGRNA和蛋白质相互作用(a)(a)匹配的SPCAS9和(b)MM5-SPCAS9聚焦HNH催化位点和PAM(NGG)区域。(C&D)显示了匹配的和MM5的不同视图,从而缩放了PAM远端和RUVC区域相互作用。T-DNA,NT-DNA和SGRNA分别为颜色的洋红色,黄色和浅蓝色。SPCAS9,HNH和RUVC的两个核酸酶结构域以白色和深蓝色显示。