XiaoMi-AI文件搜索系统
World File Search SystemsgRNA
基因编辑
将打靶特定人源基因的 Cas9 和 sgRNA 转染到 HEK293 细胞。转染所用的质粒 DNA 上含有 表达带双端核定位序列 ( NLS )的 Cas9 及 sgRNA 的表达框,通过 TransIT-X2 (Mirus) 转染 试剂进行转染。转染所用的 Cas9 mRNA 进行了假尿苷和 5- 甲基胞嘧啶修饰且带有双端 核定位序列,使用 transIT-mRNA 转染试剂将 sgRNA 和 mRNA 共转染。 Cas9 RNPs 使用脂质 体 RNAiMAX ( Life Technologies ) 进行反向转染, RNP 的终浓度为 10 nmol 。 Cas9 蛋白上不含 核定位序列。 EnGen Cas9 含有双端核定位序列。编辑效率通过 T7E1 实验进行分析,结果 以修饰百分比进行统计。
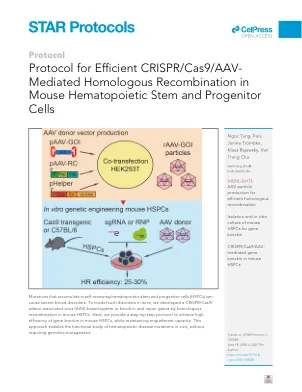
高效 CRISPR/Cas9/AAV 协议
1. 为了在小鼠 HSPC 中实现有效的同源重组 (HR) 事件,需要具有高编辑效率的特定单向导 RNA (sgRNA)。我们使用 CrispRGold 程序 ( https://crisprgold.mdc-berlin.de ) 来设计特定的 sgRNA 并预测潜在的脱靶 ( Chu et al., 2016a )。每个目标序列应设计几个特定的 sgRNA。必须通过使用 T7 内切酶 I 测定 ( Guschin et al., 2010 ) 测量错配的 DNA 异源双链体以及对至少 2 种主要血细胞类型(例如 B 细胞和 T 细胞)的 PCR 产物进行 Sanger 测序来验证所有 sgRNA 的编辑效率。可以从 IDT、Synthego 或其他供应商处订购化学修饰或未修饰形式的 sgRNA。 2. 供体模板的最佳设计对于小鼠 HSPC 中的高效 HR 至关重要。供体模板包括 5'、3' 同源臂和所需的修饰基因序列。同源臂的长度取决于目标序列的特异性,每个同源臂由 600 到 2000 bp 组成。AAV 基因组的包装能力是设计供体模板的一个限制,因为基于 AAV 的供体模板的最大长度不应超过 4.5kb。如果没有使用报告基因,则应通过引入可用于量化 HR 效率的沉默突变将限制性酶识别位点添加到修饰的基因序列中。3. 为了通过 PCR 扩增和测序量化目标位点中的 HR 和非同源末端连接 (NHEJ) 事件,必须在外部设计正向或反向引物,或两者

用于超低剂量 CRISPR/Cas9 基因编辑的脂异肽复合物
评估了含有琥珀酰四乙烯五胺 (Stp) 和脂氨基脂肪酸 (LAF) 的双 pH 响应异种肽载体用于基于 CRISPR/Cas9 的基因组编辑。使用三种不同的基因组靶标(Pcsk9、eGFP、mdx 外显子 23),在三种不同的报告细胞系中筛选了不同的载体拓扑结构、LAF/Stp 比率的变化和 LAF 类型作为 Cas9 mRNA/sgRNA 多聚复合物。鉴定出一种 U 形和三种束 (B2) 形脂异种肽,它们表现出显著的效率。在亚纳摩尔 EC 50 浓度分别为 0.4 nM sgRNA 和 0.1 nM sgRNA 的顶级 U 形和顶级 B2 载体中,即使在全 (≥ 90%) 血清中孵育后,仍观察到顶级载体的基因组编辑效力。多聚复合物与单链 DNA 模板共同递送 Cas9 mRNA/sgRNA,用于同源性定向基因编辑,导致报告细胞中 eGFP 转化为 BFP 的比例高达 38%。顶部载体被配制成多聚复合物或脂质纳米颗粒 (LNP),随后用于体内给药。制剂在 4 ◦ C 下储存时表现出长期的物理化学和功能稳定性。重要的是,静脉内注射多聚复合物或 LNP 介导肌营养不良蛋白基因的体内编辑,触发肌营养不良蛋白表达的心肌、骨骼肌和脑组织中 mRNA 外显子 23 剪接调节。

IL-17RD 慢病毒激活颗粒 (m):sc-431369-LAC
成簇的规律间隔短回文重复序列 (CRISPR) 和 CRISPR 相关蛋白 (Cas9) 系统是一种适应性免疫反应防御机制,古细菌和细菌利用该机制来降解外来遗传物质。该机制可以重新用于其他功能,包括哺乳动物系统的基因组工程,例如基因敲除 (KO) (1,2) 和基因激活 (3-5)。CRISPR 激活质粒产品利用与 VP64 激活域融合的 D10A 和 N863A 失活 Cas9 (dCas9) 核酸酶与 sgRNA (MS2) 结合,从而实现特定基因的识别和上调,sgRNA (MS2) 是一种靶向特异性 sgRNA,经过设计可结合 MS2-P65-HSF1 融合蛋白 (5)。这种协同激活介质 (SAM) 转录激活系统* 提供了一个强大的系统,可最大限度地激活内源性基因表达 (5)。

用于分析汇集的 CRISPR 筛选的算法基准
简介 成簇的规则间隔回文重复序列 (CRISPR) 和 CRISPR 相关 (Cas) 蛋白是一类由细菌编码的、RNA 引导的可编程 DNA 靶向和切割系统。由于其使用可定制的单向导 RNA (sgRNA) 的可编程特性,CRISPR-Cas 已实现强大的汇集筛选,以探索基因组范围内遗传扰动的功能。以最常用的 CRISPR-Cas9 系统为例,化脓性链球菌 Cas9 蛋白可以与 110 个核苷酸 (nt) 的 sgRNA 复合,该 sgRNA 包含一个 20 nt 序列,该序列与目标 DNA 区域互补结合并诱导双链断裂 (DSB)。基因组 DNA 上的这种切割机制会触发宿主非同源末端连接 (NHEJ) 或同源定向修复 (HDR)

Nyberg 等人 BioRxiv
通过同源定向修复 (HDR) 进行基因组编辑使得对基因序列进行精确而慎重的修改成为可能。CRISPR/Cas9 介导的 HDR 是实现这一目标的最简单方法。然而,在提高效率和扩大对果蝇以及其他果蝇物种的任何遗传背景的适用性方面仍然存在技术挑战。为了解决这些问题,我们开发了一种两阶段标记辅助策略,以促进果蝇的精确、无疤痕编辑,而几乎不需要分子筛选。使用与重组 Cas9 蛋白复合的 sgRNA,我们分析了每个 sgRNA 的基因组切割效率。然后,我们使用有效切割目标基因的 sgRNA 和转化标记的新应用进行 HDR。这些新工具可用于在感兴趣的区域进行单个更改或一系列等位基因替换,或创建其他遗传工具,例如平衡染色体。
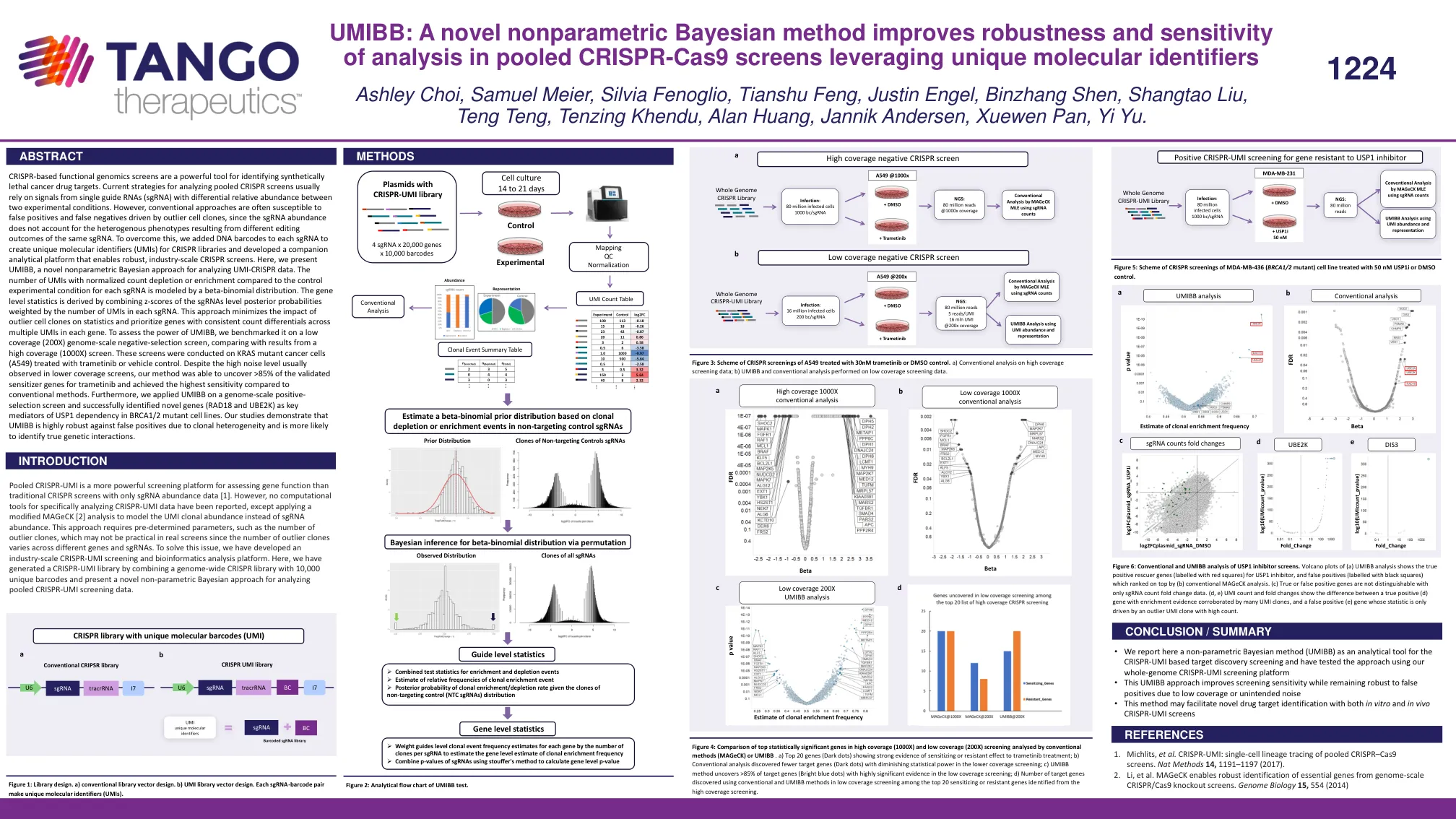
UMIBB:一种新的非参数贝叶斯方法提高了稳健性和灵敏度
基于 CRISPR 的功能基因组学筛选是识别合成致死癌症药物靶点的有力工具。目前分析汇集的 CRISPR 筛选的策略通常依赖于来自在两种实验条件下具有不同相对丰度的单个向导 RNA (sgRNA) 的信号。然而,传统方法通常容易受到由异常细胞克隆驱动的假阳性和假阴性的影响,因为 sgRNA 丰度不能解释由相同 sgRNA 的不同编辑结果导致的异质表型。为了克服这个问题,我们在每个 sgRNA 中添加了 DNA 条形码,以创建 CRISPR 文库的唯一分子标识符 (UMI),并开发了一个配套的分析平台,以实现强大的工业规模 CRISPR 筛选。在这里,我们介绍了 UMIBB,一种用于分析 UMI-CRISPR 数据的新型非参数贝叶斯方法。与每个 sgRNA 的对照实验条件相比,具有标准化计数消耗或富集的 UMI 数量由 beta-二项分布建模。基因水平统计数据是通过将 sgRNA 水平后验概率的 z 分数与每个 sgRNA 中 UMI 的数量加权而得出的。这种方法最大限度地减少了异常细胞克隆对统计数据的影响,并优先考虑每个基因中多个 UMI 之间计数差异一致的基因。为了评估 UMIBB 的功效,我们在低覆盖率(200X)基因组规模负选择筛选上对其进行了基准测试,并与高覆盖率(1000X)筛选的结果进行了比较。这些筛选是在用曲美替尼或载体对照处理的 KRAS 突变癌细胞(A549)上进行的。尽管在较低覆盖率筛选中通常会观察到高噪音水平,但我们的方法能够发现 >85% 的曲美替尼已验证的致敏基因,并且与传统方法相比实现了最高的灵敏度。此外,我们将 UMIBB 应用于基因组规模的正向选择筛选,并成功确定了新基因(RAD18 和 UBE2K)是 BRCA1/2 突变细胞系中 USP1 依赖性的关键介质。我们的研究表明,UMIBB 对克隆异质性导致的假阳性具有很高的稳健性,并且更有可能识别真正的遗传相互作用。

CRISPR稳定的敲入细胞生产(CAT。C.C408)案例研究:使用CRISPR将红色荧光蛋白(RFP)基因敲入人类胚胎
图2 PCAS-GUIDE-AAVS1和PAAVS1-RFP-DNR的矢量图。pCAS指向AAVS1是哺乳动物细胞中SGRNA和Cas9共表达的多合一载体。SGRNA的表达是由强大的组成型Pol III启动子U6启动子驱动的。而CMV启动子则驱动CAS9酶的表达。paAVS1-RFP-DNR在CMV启动子下的PGK启动子和RFP基因下表达紫霉素的抗性标记。5'和3'AAVS1同源臂(“ aavs-right”和“ aavs-left”)为单元提供了一个用于同源性修复的模板。

基因敲除试剂盒
我们强烈建议您在使用靶向特异性多向导 sgRNA 之前,先使用特定细胞类型的阳性对照优化转染条件。为了优化人类细胞系的条件,EditCo 提供了转染优化试剂盒,其中包含靶向人类 TRAC 的阳性对照多向导 sgRNA。对于小鼠细胞系,我们建议使用靶向小鼠 Rosa26 的阳性对照单向导 sgRNA 来优化您的条件(请参阅第 2 页所需的其他材料)。我们建议为您的基因组编辑实验形成核糖核蛋白 (RNP) 复合物,以最大限度地提高编辑效率并减少脱靶效应。选择 EditCo 的电穿孔、脂质转染或核转染方案与此试剂盒一起使用。所有方案均可在 editco.com/resources 上找到。

INSL5 CRISPR 激活质粒 (h): sc-417448-ACT
成簇的规律间隔短回文重复序列 (CRISPR) 和 CRISPR 相关蛋白 (Cas9) 系统是一种适应性免疫反应防御机制,古细菌和细菌利用该机制来降解外来遗传物质。该机制可以重新用于其他功能,包括哺乳动物系统的基因组工程,例如基因敲除 (KO) (1,2) 和基因激活 (3-6)。CRISPR 激活质粒产品利用与 VP64 激活域融合的 D10A 和 N863A 失活 Cas9 (dCas9) 核酸酶与 sgRNA (MS2) 结合,从而实现特定基因的识别和上调,sgRNA (MS2) 是一种靶向特异性 sgRNA,经过设计可结合 MS2-P65-HSF1 融合蛋白 (6)。这种协同激活介质 (SAM) 转录激活系统提供了一个强大的系统,可最大限度地激活内源性基因表达 (6)。