XiaoMi-AI文件搜索系统
World File Search SystemshRNA

通过顺序靶向肿瘤缺氧实现有效的药物和 shRNA 递送,以协同治疗三阴性乳腺癌
由于缺乏针对性的治疗方法,三阴性乳腺癌的临床治疗仍然具有挑战性。由于三阴性乳腺癌具有高度缺氧性且HIF-1α的表达高于其他亚型,我们制备了缺氧响应性聚合物胶束,共负载药物和shRNA,通过靶向缺氧肿瘤微环境,随后在缺氧条件下靶向过表达的HIF-1α来治疗三阴性乳腺癌。胶束由甲氧基聚乙二醇(mPEG)和聚-L-赖氨酸(PLL)共聚物组成,以AZO作为mPEG和PLL之间的缺氧响应桥。一旦暴露于缺氧,AZO桥就会断裂,导致胶束解体并快速释放。体外和体内结果表明,通过对缺氧的敏感反应,胶束能够同时将药物和shRNA递送到缺氧部位并实现位点特异性快速释放;缺氧响应性shRNA递送有效沉默HIF-1α及其下游基因,不仅改善缺氧肿瘤对药物的反应,而且调节肿瘤微环境以进一步改善药物和shRNA递送;因此,化疗和HIF-1α靶向基因治疗的协同治疗在小鼠原位TNBC模型中抑制了原发性TNBC肿瘤的生长及其远处转移。缺氧响应性聚合物胶束因其良好的生物相容性而成为一种安全、有效且普遍适用的药物和基因载体,可用于治疗TNBC以及其他缺氧肿瘤。

假木香酸 B 靶向 CD147 选择性杀死急性...
表 3-1. CD147 SHRNA 序列...................................................................................................................... 28
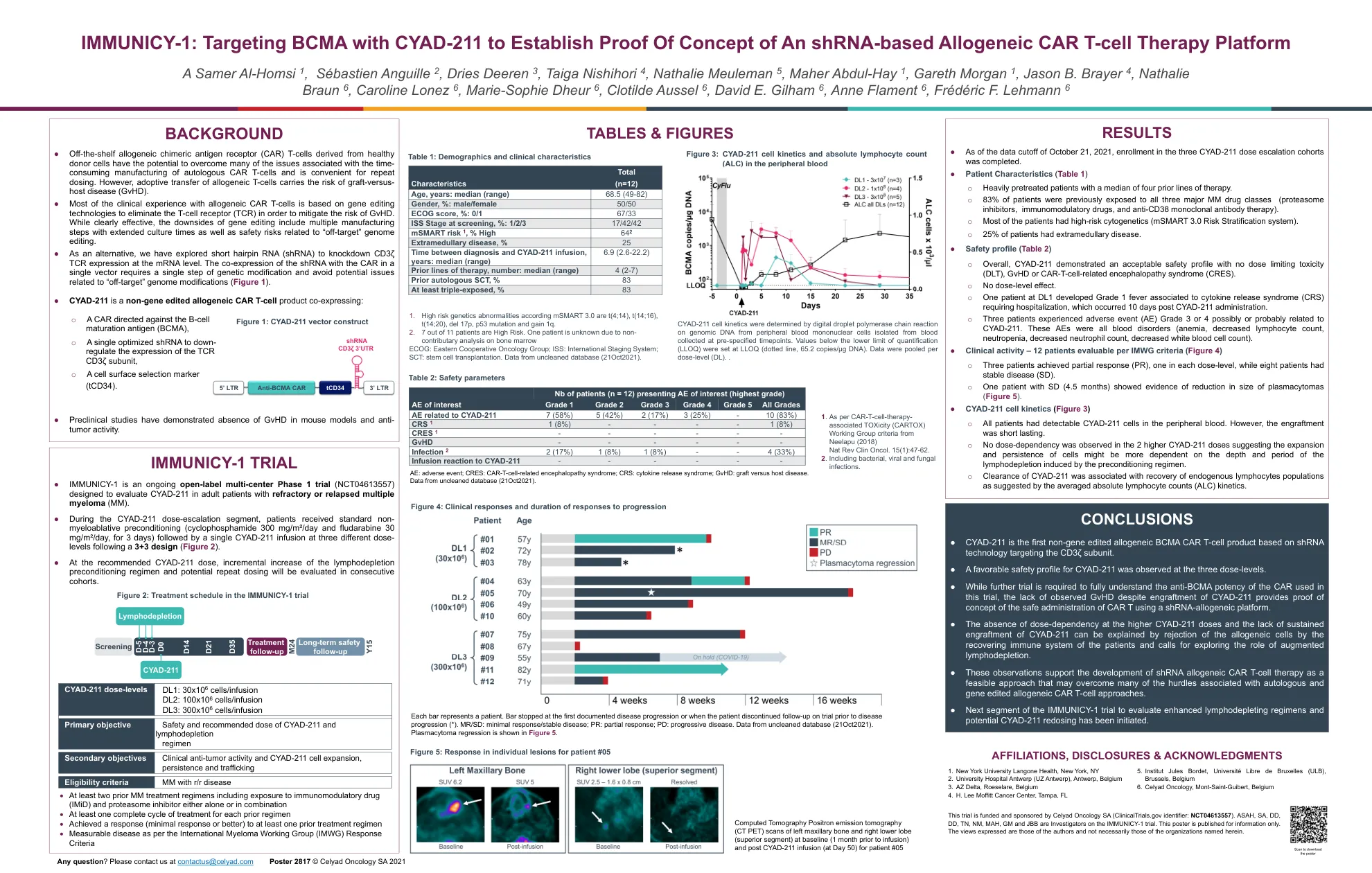
IMMUNICY-1:利用 CYAD-211 靶向 BCMA,建立基于 shRNA 的同种异体 CAR-T 细胞治疗平台的概念验证
● 源自健康供体细胞的现成同种异体嵌合抗原受体 (CAR) T 细胞有可能克服自体 CAR T 细胞制造耗时带来的许多问题,并且便于重复给药。然而,同种异体 T 细胞的过继转移存在移植物抗宿主病 (GvHD) 的风险。
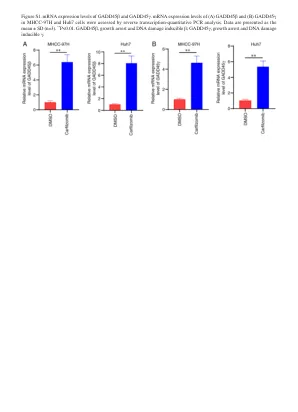
mRNA的表观遗传调节介导表型...
图S2。 通过蛋白质印迹评估的GADD45αshRNA的沉默效率。 gADD45α蛋白表达水平在(a)MHCC -97H和(b)用NC和三个靶向GADD45α的SHRNA后的HUH7细胞中。 基于GADD45α蛋白的表达,SH2的沉默是最重要的,用于随后的实验。 数据表示为平均值±SD(n = 3)。 ** p <0.01。 GADD45α,生长停滞和DNA损伤诱导α; NC,阴性对照; SH,短发夹。图S2。通过蛋白质印迹评估的GADD45αshRNA的沉默效率。gADD45α蛋白表达水平在(a)MHCC -97H和(b)用NC和三个靶向GADD45α的SHRNA后的HUH7细胞中。基于GADD45α蛋白的表达,SH2的沉默是最重要的,用于随后的实验。数据表示为平均值±SD(n = 3)。** p <0.01。GADD45α,生长停滞和DNA损伤诱导α; NC,阴性对照; SH,短发夹。

Colt-Cancer
说明:柯尔特 - 癌数据库是SHRNA辍学特征的集合,涵盖了约16000个人类基因,并使用在COLT(CCBR-OICR LINTIVIRAL TECHECTION)FATILION中开发的微阵列检测平台源自70多个胰腺,卵巢和乳腺癌细胞线。所有SHRNA辍学配置文件均可以通过此网站免费获得或查询。
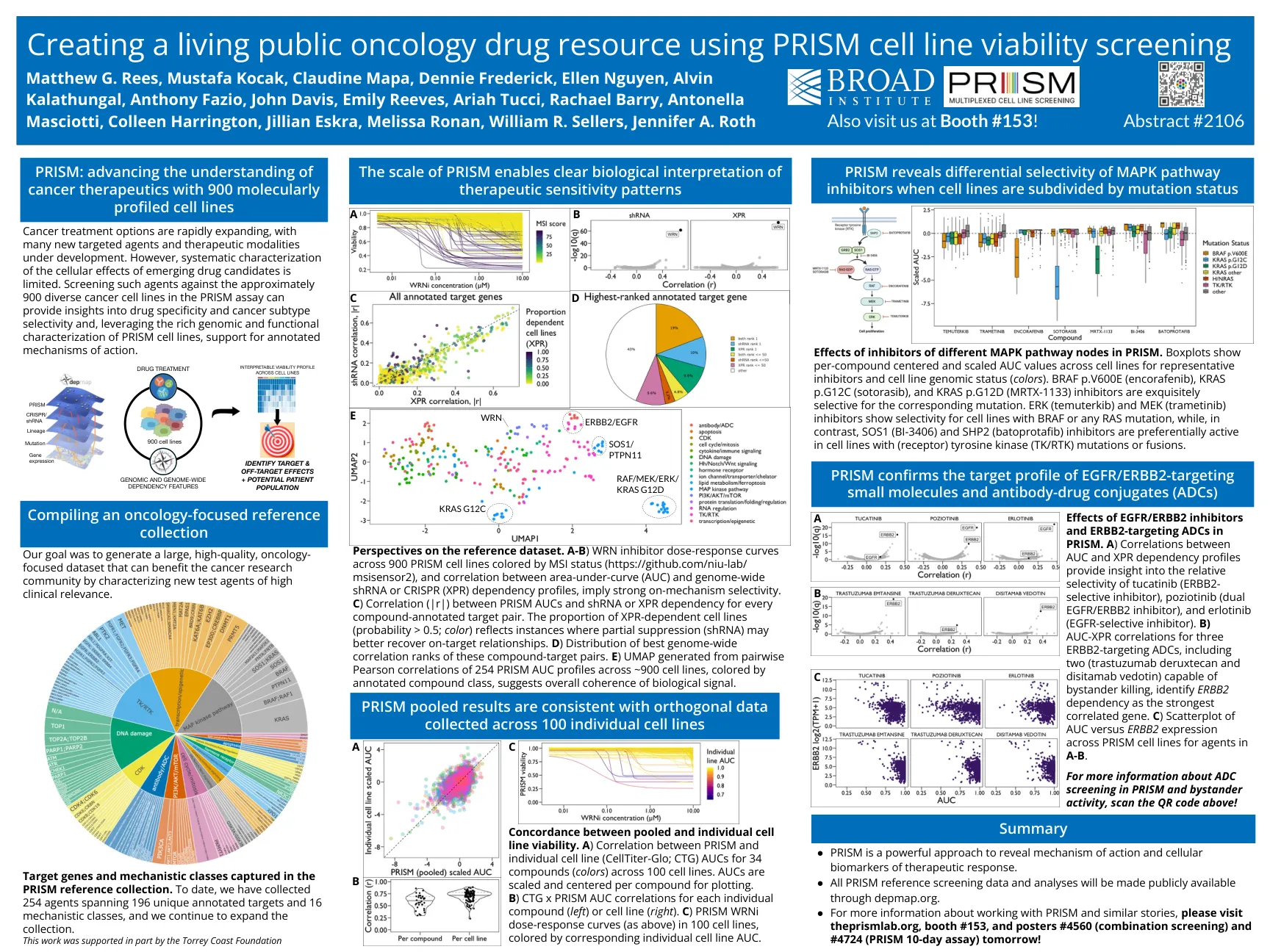
2024 ONCREF AACR海报
参考数据集的观点。a-b)WRN抑制剂剂量反应曲线横跨900个Prism细胞系(https://github.com/niu-lab/ msisensor2),以及区域范围内curve(auc)和基因组 - 含量 - 含基因组shrna或crispr(xpr)依赖性(xpr)依赖性的相关性,依赖于强度,是强化的良好。c)每个化合物注销的目标对之间的棱镜AUC和shRNA或XPR依赖性之间的相关性(| r |)。XPR依赖性细胞系的比例(概率> 0.5;颜色)反映了部分抑制(SHRNA)可以更好地恢复目标上的关系的实例。d)这些化合物靶向对的最佳全基因组相关等级的分布。e)由254个Prism AUC auc pro填充的成对Pearson相关性产生的UMAP在约900个细胞系中,通过注释化合物颜色,表明生物学信号的总体连贯性。
发夹和剪刀 - 为大众提供非基因编辑的同种异体 CAR-T 细胞疗法 David Gilham Celyad Oncology 首席科学官
概述 两种嵌合抗原受体 (CAR) T 细胞疗法获批用于治疗 B 细胞恶性肿瘤,凸显了细胞免疫疗法在提供令人印象深刻的持久临床反应方面的潜力 1 。这些产品本质上是自体的,涉及从患者身上收集用于制造 CAR T 细胞的免疫细胞。一旦生产出来,这些 CAR T 细胞就会作为临床产品重新注入患者体内。然而,自体疗法面临着重大挑战,包括产品生产时间(目前需要数周),在此期间患者的病情可能会恶化,以及起始材料的质量高度不稳定,这可能导致制造失败。同种异体 CAR T 细胞疗法是一种现成的方法,可以在需要时进行管理,是理想的解决方案。这种方法从健康供体中生成细胞,形成一个 CAR T 细胞库,可根据需要使用。同种异体 CAR T 的关键挑战是克服与同种异体 CAR T 细胞识别健康患者组织相关的毒性。这是由 T 细胞受体 (TCR) 介导的。破坏 TCR 是所有当前同种异体 CAR-T 策略的基础 2 。发夹和剪刀目前,用于生成同种异体 CAR-T 的基因编辑技术处于临床开发的早期阶段。不同的基因编辑方法都是基于切割编码 TCR 的基因之一内的基因组,从而永久性地降低整个 TCR 复合物的表达。虽然是一种优雅的方法,但由于潜在的产品安全问题,这种剪刀策略一直难以进入临床测试阶段——主要是确保在基因编辑过程中没有“脱靶”基因组切割 3 。或者,在 mRNA 水平上靶向基因表达不涉及切割基因组,并避免危及基因组完整性。为了实现这种 mRNA“编辑”,Celyad Oncology 采用了短发夹 RNA (shRNA),这是一种几十年来用于敲低基因表达的方法 4 。该方法涉及使用具有与目标基因互补序列的 shRNA。换句话说,靶向 shRNA 可以通过干扰 mRNA 而不是切割基因组 5 来特异性降低所需蛋白质(如 TCR 复合物)的水平。其中的核心是一体化载体方法。只需一步,将单一试剂(载体)引入健康供体 T 细胞,即可同时产生 T 细胞中的所有元素,这些元素可以将 T 细胞重定向到肿瘤(CAR)、消除 TCR(shRNA)并提供一个手柄,使修饰的细胞可以在制造过程中富集(标记物)。同种异体 CAR T 细胞平台中的 shRNA CD3z 亚基为 TCR 提供主要信号功率,从而激活和参与 T 细胞杀伤能力。通过选择最佳 shRNA 和工艺开发,靶向 CD3z 可使原代 T 细胞上的 TCR 持续高水平敲低,达到与基因编辑 CD3z 基因时相同的水平(图 1A)。从功能上讲,这与这些细胞无法对有丝分裂刺激(又称 TCR 驱动的 T 细胞活化;图 1B)作出反应以及当这些细胞被注入黄金标准体内测试模型时相应没有毒性有关(图 2A、B)。有趣的是,shRNA 靶向 T 细胞的持久性比 CRISPR-Cas9 基因的持久性要长得多

摘要PDF海报
摘要:细胞类型之间的转分化依赖于基于知识的搜索最佳重编程因素。我们最近的研究发现,ASCL1,MiR9/9*-124,NPTB shRNA和p53 shRNA的过表达有效地将人皮肤成纤维细胞转换为神经元。通过分析人类皮肤成纤维细胞的纵向RNA-seq数据通过这些重编程因子的各种组合进行转化,我们构建了基因调节网络(GRN)模型,捕获了对神经元转化重要的高阶信息。GRN中基因群落和转录因子(TF)的检查确定OTX2和LMX1A是转化为神经元的关键调节剂,因为它们与与神经元发育和分化功能相关的基因的连接最强。 我们通过实验证实了OTX2和LMX1A的关键作用,因为它们的敲低显着损害了转换。 研究表明,GRN模型有效地扩大了人类皮肤成纤维细胞转差为神经元的经验发现最佳重编程因子。 这种方法的进一步改进可以确定直接细胞转换的普遍适用原理。GRN中基因群落和转录因子(TF)的检查确定OTX2和LMX1A是转化为神经元的关键调节剂,因为它们与与神经元发育和分化功能相关的基因的连接最强。我们通过实验证实了OTX2和LMX1A的关键作用,因为它们的敲低显着损害了转换。研究表明,GRN模型有效地扩大了人类皮肤成纤维细胞转差为神经元的经验发现最佳重编程因子。这种方法的进一步改进可以确定直接细胞转换的普遍适用原理。

Prism合作者幻灯片的资源页面
●针对MTA合并性PRMT5抑制剂(在第1/2阶段试验中)的基因表达,蛋白质组学,shRNA敲低和广泛的重新利用数据集相关性,据报道优先针对MTAP-MTAP-浸没细胞活性。●分别具有PRMT5和MTAP的化合物靶标和预期的生物标志物是最强的命中率之一,与Prism中的复合响应相关

人胎盘中合成肌细胞细胞谱系发育的关键调节剂
背景。胎盘是一种瞬态器官,在怀孕期间形成以支持胎儿发育并调节影响慢性疾病风险的环境线索的暴露。胎盘在许多方面支持胎儿发育,包括促进营养和氧气交换,去除有害废物产品,产生关键的激素(例如人类绒毛膜促性腺激素)以及提供免疫保护。这些功能在很大程度上是由被称为合胞素细胞和额外滋养细胞细胞的终末分化的滋养细胞执行的。尽管合成肌细胞细胞和跨性滋养细胞细胞的重要性,但仍不清楚它们如何专门支持最佳胎儿发育。目标。使用功能方法丧失来确定合成肌细胞细胞谱系发育的转录调节因子。方法。候选转录因子(TBX3,VGLL3和ATF3)使用慢病毒介导的短发蛋白RNA(SHTBX3,SHTBX3,SHVGLL3或SHATF3)使用胞质衍生的人滋养细胞干细胞中击倒。将非特异性shRNA(SHCONTROL)用作对照。转导后,使用紫霉素选择细胞,并分别通过RT-QPCR和Western印迹在转录本和蛋白质水平上确认敲低效率。通过功能和转录组评估评估了转录因子敲低对滋养细胞干细胞分化为合成型肉芽细胞的影响。结果。结论。未来的方向。与用SHControl转导的细胞相比,SHTBX3和SHVGLL3的转导在合成型细胞细胞分化后导致形态异常。 可以使用滋养细胞干细胞中的功能方法丧失来评估候选转录调节剂对合成细胞细胞谱系发育的关键贡献。 初步结果表明,TBX3和VGLL3对于建立合成型细胞细胞谱系至关重要。 然而,需要更深入的表征来识别TBX3和VGLL3调节合成细胞成分的发育的分子机制。 未来的研究将包括完成剩余的候选转录因子,ATF3,全基因组评估(例如ATAC-SEQ)的shRNA敲低,以及所有SHRNA转换的其他功能输出,例如人类绒毛膜促性腺激素的产生。在合成型细胞细胞分化后导致形态异常。可以使用滋养细胞干细胞中的功能方法丧失来评估候选转录调节剂对合成细胞细胞谱系发育的关键贡献。初步结果表明,TBX3和VGLL3对于建立合成型细胞细胞谱系至关重要。然而,需要更深入的表征来识别TBX3和VGLL3调节合成细胞成分的发育的分子机制。未来的研究将包括完成剩余的候选转录因子,ATF3,全基因组评估(例如ATAC-SEQ)的shRNA敲低,以及所有SHRNA转换的其他功能输出,例如人类绒毛膜促性腺激素的产生。