XiaoMi-AI文件搜索系统
World File Search Systemstim1
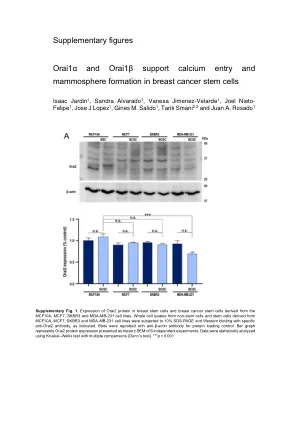
补充数字ORAI1α和Orai1β支持钙...
补充图2。源自WT MDA-MB-231和ORAI1-KO MDA-MB-231的乳腺癌干细胞(BCSC)中Orai2,Orai3,stim1和stim2的表达。从WT和Orai1-KO(O1KO)MDA-MB-231细胞得出的干细胞中的全细胞裂解物进行10%SDS-PAGE,并用特异性抗ORAI2,抗ORAI2,抗Orai3,anti-Orai3,anti-stim1和anti-stim1和anti-2抗体进行蛋白质印迹。印迹。bar图代表Orai2(a),Orai3(b),STIM1(C)和Stim2(D)蛋白表达,以4个单独的实验的平均值±SEM表示。使用Mann-Whitney U检验对数据进行了统计分析。**** p <0.0001

ORAI1抑制作肾小管肌病和Stormorken综合征的有效临床前疗法
引言管状肌病(TAM)是一种罕见的肌肉障碍,具有广泛的表型异质性,从无症状高度高血症到渐进的儿童期形式,具有严重的上肢和下肢肌肉无力,肌痛,肌痛,抽筋和膜(1,2)。大多数患者都表现出其他多系统的体征,例如MIOSIS,血小板减少症,次生症,鱼质病,身材矮小和阅读障碍,并且完整的临床图片称为Stormorken综合征(Strmk)(Strmk)(2-4)。tam和strmk(Omim#160565和#615883)形成临床连续性,病理学迹象的发生和程度取决于所涉及的基因和突变的位置。大多数TAM/Strmk患者在Stim1中携带杂合的错义突变,编码居住在内质/肌浆网(ER/SR)的无处不在表达的Ca 2+传感器。在Ca 2+引起的EF手臂中的突变基本上会导致肌肉表型,偶尔会导致血小板和皮肤,脾脏,骨骼和骨骼的异常耗尽(5-13),而胞质盘旋型域的突变则与coiled-Coiled-Coil 304残基相关(均具有10个残留型),并具有104个杂物(均为804的杂物)。 14-19)。等离子体内置的Ca 2+通道中的错义突变不太普遍,要么影响孔形成的跨膜结构域,并引起严重的表型,并具有明显的肌肉无力,Miosis,出血性核糖症,鱼囊和

ORAI1/stims肺血管疾病中的调节剂
钙(Ca 2 +)是调节各种细胞过程的次级信使。但是,CA 2 +不当行为可能导致病理状况。ORAI1是CA 2 +通道,有助于商店经营的钙进入(SOCE),并且在几种细胞类型的Ca 2 +稳态中起着至关重要的作用。ORAI1的失调导致严重的免疫缺陷综合征,一些癌症,肺动脉高压(PAH)和其他心肺疾病。 在其激活过程中,Orai1主要由基质相互作用分子(STIM)蛋白,尤其是stim1调节。但是,最近还描述了许多其他监管伙伴。 对这些调节伙伴的知识越来越多,可以更好地了解SOCE的下游信号传导途径,并提供了一个绝佳的机会,可以使Orai1失调在这些disse中解密。 这些蛋白质参与其他细胞功能,使其具有吸引力的治疗靶标。 本综述主要关注ORAI1调节伙伴在肺循环和炎症的生理和病理条件下。导致严重的免疫缺陷综合征,一些癌症,肺动脉高压(PAH)和其他心肺疾病。在其激活过程中,Orai1主要由基质相互作用分子(STIM)蛋白,尤其是stim1调节。但是,最近还描述了许多其他监管伙伴。对这些调节伙伴的知识越来越多,可以更好地了解SOCE的下游信号传导途径,并提供了一个绝佳的机会,可以使Orai1失调在这些disse中解密。这些蛋白质参与其他细胞功能,使其具有吸引力的治疗靶标。本综述主要关注ORAI1调节伙伴在肺循环和炎症的生理和病理条件下。
8688医疗多重
Congenital myopathy / Congenital muscular dystrophy (COL6A1, COL6A2, COL6A3, COL12A1, FKRP, FKTN, LAMA2, LARGE1, POMGNT1, POMGNT2 (GTDC2), POMT1, POMT2, COL4A1, COL4A2, DAG1, DPM1, DPM2, DPM3, dolk, ISPD, GMPPB, b3galnt2, chkb, plec, sil1, b4gat1 (b3gnt1), pomk (sgk196), itga7, Them5, Micu1, act1, cfl2, dnm2, tbd1, mbt1, mbt1, mbt1, myh8,neb,ryr1,sepn1,tnni2,tnnt1,tnnt3,tpm2,tpm3,stim1,ecel1,cdc78,kbtbd10(kbtbd10),klhl40(kbtbd5)(kbtbd5),mybdbd),mybdbd),mybd) ),mybd),mybd),mybd) Lamp2, VMA21, STAC3, lmod3, MEGF10, epg5, ttn, adamts15, cacna1s, CNTN1, Doc7, Golga2, Hacd1 (PLPLA), inpp5k, klhl9, msto1, Mtm18, mybpp, mybpp, mybpp, mybpp, mybpp srpk3,them38a,trappc11)

神经系统中的钙池操纵钙通道
钙库操纵的钙离子内流 (SOCE) 是一种广泛的细胞钙离子信号传导机制,它源于钙离子通过 Orai 家族钙通道跨质膜流入,以响应细胞内钙离子库的消耗。Orai 通道是神经元和神经胶质细胞中一种重要的钙离子内流机制,它由一种独特的由内而外的门控过程激活,该过程涉及与内质网钙离子传感器 STIM1 和 STIM2 的相互作用。最近的证据表明,SOCE 广泛存在于神经系统的所有领域,而它的生理学和病理生理学现在才刚刚开始被人们了解。在这里,我们回顾了有关神经系统中 SOCE 机制及其对基因表达、神经元兴奋性、突触可塑性和行为的贡献的越来越多的文献。我们还探讨了 SOCE 与神经系统疾病之间日益密切的联系,并讨论了针对 SOCE 对脑部疾病的治疗意义。

论文模板
ATP ATP腺苷-5'-三磷酸凸轮钙调蛋白CARQ CAQ+激活的Rho蛋白,带有嵌入的IQP Ceru ceru cerulean,相当于CFP CFP CFP CyAn荧光蛋白 Dulbecco's modified eagle medium FBS Fetal Bovine Serum FKBP12 12-kDa FK506 and rapamycin-binding protein FRB FKBP-rapamycin binding domain FRET Fluorescence resonance energy transfer GST Glutathione S-transferase His Polyhistidine-tag IRES Internal ribosomal entry site LB Luria Broth LOV Light-oxygen-voltage域,lov2域Lovs1K Lov2结构域与刺激1 c末端碎片MCS多个克隆位点MLCKP肌球蛋白轻链激酶激酶肽MRFP单体红色荧光蛋白相当于RFP,相当于RFP NES核出口NLS NLS NLS信号NLS信号NLS核定位PBS PBS PBS磷酸盐磷酸盐磷酸盐磷酸盐磷酸盐磷酸盐磷酸盐酶磷酸盐磷酸盐磷酸盐磷酸盐磷酸盐磷酸盐磷酸盐磷酸盐磷酸盐反应pdbu pdbu pdbu pdbu pdbu pdbu pdbu pdbu pdbu pdbu pdbu pdbu pdbu pdbu pdbu pdbu pdbu pdbu pdbu pdbu PKC Protein kinase C pLyn Palmitoylation sequence of Lyn kinase RFP Red fluorescent protein, equivalent to mRFP SDS-PAGE Sodium dodecyl sulfate polyacrylamide gel electrophoresis SH3 SRC Homology 3 Domain TEV Tobacco etch virus TEVp Tobacco etch virus protease TS Temperature-sensitive tsTEVp Temperature-sensitive tobacco蚀刻病毒蛋白酶tvmvp烟草静脉斑点病毒蛋白酶蛋白酶ven venus,相当于YFP YFP YFP黄色荧光蛋白,相当于Ven

出版物 Liebing AD、Rabe P、Krumbholz P、Zieschang C、Bischof F、Schulz A、Billig S、Birkemeyer C、Pillaiyar T、Garcia- Marcos M、Kraft R、Stäu
出版物 Liebing AD, Rabe P, Krumbholz P, Zieschang C, Bischof F, Schulz A, Billig S, Birkemeyer C, Pillaiyar T, Garcia-Marcos M, Kraft R, Stäubert C (2025) 琥珀酸受体 1 信号转导相互依赖于亚细胞定位和细胞代谢。 FEBS J doi:10.1111/febs.17407 Röthe J, Kraft R , Ricken A, Kaczmarek I, Matz-Soja M, Winter K, Dietzsch AN, Buchold J, Ludwig MG, Liebscher I, Schöneberg T, Thor D (2024) 小鼠粘附 GPCR GPR116/ADGRF5 在胰岛调节中具有双重功能生长抑素释放和胰岛发育。共同生物学7:104。 Kaczmarek I、Wower I、Ettig K、Kuhn C、Kraft R、Landgraf K、Körner A、Schöneberg T、Horn S、Thor D (2023) 使用创新的 RNA-seq 数据库 FATTLAS 识别参与脂肪组织功能的 GPCR。iScience 26:107841。Peters A、Rabe P、Liebing AD、Krumbholz P、Nordström A、Jäger E、Kraft R、Stäubert C (2022) 羟基羧酸受体 3 和 GPR84 – 两种在先天免疫细胞中具有相反功能的代谢物感应 G 蛋白偶联受体。Pharmacol Res 176:106047。 Rabe P、Liebing AD、Krumbholz P、Kraft R、Stäubert C (2022) 琥珀酸受体 1 抑制对谷氨酰胺上瘾的癌细胞的线粒体呼吸。Cancer Lett 526:91-102。Peters A、Rabe P、Krumbholz P、Kalwa H、Kraft R、Schöneberg T、Stäubert C (2020) 羟基羧酸受体 3 和 G 蛋白偶联受体 84 的自然偏向信号传导。Cell Commun Signal 18:31。Röthe J、Kraft R、Schöneberg T、Thor D (2020) 探索原发性胰腺胰岛中的 G 蛋白偶联受体信号传导。Biol Proced Online 22:4。 Stegner D, Hofmann S, Schuhmann MK, Kraft P, Herrmann AM, Popp S, Höhn M, Popp M, Klaus V, Post A, Kleinschnitz C, Braun A, Meuth SG, Lesch KP, Stoll G, Kraft* R , Nieswandt* B (2019) Orai2 介导的电容性 Ca 2+ 条目的丢失具有神经保护作用急性缺血性中风。笔画 50:3238-3245。 Röthe* J、Thor* D、Winkler J、Knierim AB、Binder C、Huth S、Kraft R、Rothemund S、Schöneberg T、Prömel S (2019) 粘附 GPCR 卵白蛋白参与调节胰岛素释放。 Cell Rep 26:1573-1584。Kraft R (2015) 神经系统中的 STIM 和 ORAI 蛋白。Channels (Austin) 9:235-243。Michaelis M、Nieswandt B、Stegner D、Eilers J、Kraft R (2015) STIM1、STIM2 和 Orai1 调节钙池操纵的钙内流和小胶质细胞的嘌呤能激活。Glia 63:652-663。Kallendrusch S、Kremzow S、Nowicki M、Grabiec U、Winkelmann R、Benz A、Kraft R、Bechmann I、Dehghani F、Koch M (2013) G 蛋白偶联受体 55 配体 L-α-溶血磷脂酰肌醇在兴奋毒性损伤后发挥小胶质细胞依赖性神经保护作用。 Glia 61:1822-1831。Wegner F、Kraft R、Busse K、Härtig W、Leffler A、Dengler R、Schwarz J(2012 年)分化的人类中脑衍生神经祖细胞表达含有 α2β 亚基的兴奋性士的宁敏感甘氨酸受体。PLoS One 7:e36946。