XiaoMi-AI文件搜索系统
World File Search Systemstrand

编码复合DNA以纠正替换,链损失和缺失
DNA分子上的数据存储是存档大量数据的有前途的方法[1] - [4]。在经典的DNA存储系统中,将二进制信息编码为由四个DNA碱基{a,c,g,t}组成的序列。编码序列用于使用DNA合成的生化过程生成称为链的DNA分子。合成的链储存在管中。要检索二进制信息,必须通过DNA测序读取链,并将解码回到二进制表示中。合成过程和测序程序是容易出错的,并且随着DNA的自然降解,它们会向DNA链引入错误。为了确保数据可靠性,必须通过算法和错误校正代码(ECC)来纠正错误。最近,为了允许更高的潜在信息能力[5],[6]引入了复合DNA合成方法。在此方法中,使用标准DNA合成方法创建的多个副本可用于创建复合DNA符号,该符号由DNA碱基的混合物及其比率定义,其比率及其特定位置。通过定义不同的混合物和比率,可以将字母扩展到具有4个以上的符号。更正式地,可以将特定位置的复合DNA符号抽象为概率的四重奏{p a,p c,p g,p g,p t},其中p x,0≤px≤1是基本x∈{a,c,g,t}的底数。因此,要识别复合符号,需要对多个读数进行测序,然后在每个位置估算p a,p c,p g,p t。由于该方法中字母符号的独特结构,基本级别的误差可以轻松更改观察到的碱基的混合物及其比率,因此更改了观察到的复合符号。此外,在此设置中,合成过程的固有冗余性(即,每股多个副本)不能直接用于

TOPAS-nBio 模拟温度依赖性的间接 DNA 链断裂产量
当细胞受到低 LET 辐射(60 Co 约为 0.3 keV/µm)时,大多数 DNA 损伤不是由辐射场与 DNA 的直接相互作用引起的,而是由辐解后的化学反应引起的。因此,辐射化学对于理解电离辐射造成的生物损伤的潜在机制至关重要。蒙特卡洛径迹结构 (MCTS) 代码可以详细模拟细胞等介质中的粒子径迹。几种 MCTS 代码已经进一步开发,具有模拟水的辐解和随后的非均相化学的能力。最初的 MCTS 模拟使用纯水作为目标,并叠加 DNA 几何形状来表征物理相互作用(Charlton 1986)。现在,MCTS 代码已经变得更加复杂,可以将电离辐射的物理化学过程与 DNA 几何模型相结合。

口腔癌发生发展中 DNA 双链断裂修复的研究进展
摘要:我们探讨了与 DNA 双链断裂反应和修复相关的基因缺陷导致口腔潜在恶性疾病 (OPMD) 恶性转化为口腔鳞状细胞癌 (OSCC) 的可能性。同源重组/范康尼贫血 (HR/FA) 缺陷,而非非同源末端连接缺陷,导致 DNA 修复途径似乎与易患 OSCC 的家族性疾病特征一致 (FA、布卢姆综合征、毛细血管扩张性共济失调);对于发生在年轻患者身上的 OSCC 来说也是如此,有时这些患者很少或没有接触过经典风险因素。即使在先天性角化不良症(一种也易患 OSCC 的端粒酶复合物疾病)中,维持端粒长度的尝试也涉及一条具有共享 HR 基因的途径。因此,HR/FA 途径中的缺陷似乎在易患 OSCC 的疾病中起着关键作用。还有一些证据表明,HR/FA 通路异常与偶发病例 OPMD 和 OSCC 的恶性转化有关。我们提供的数据表明,与致命细胞系相比,一系列 OPMD 衍生的永生角质形成细胞系中 HR/FA 基因以细胞周期依赖性方式过度表达。本研究中的观察结果有力地证明了 HA/FA DNA 修复通路在 OSCC 发展中的重要作用。

Strand Life Sciences启动门户,以帮助患有罕见病的医生
诊断; 2025年2月28日,为班加罗尔的低收入患者提供折扣测试:在罕见疾病日之际,Reliance Industries子公司和领先的基因组学公司Strand Life Sciences和Life Sciences,今天启动了Strandomics Portal,以改善罕见疾病的诊断。这个新的在线门户网站可让临床医生快速轻松地访问有关遗传变异的信息,从而为每种情况提供全面的视野。这意味着患者更快,更准确地诊断。除了门户网站外,Strand Life Sciences还将以降低经济不利背景的患者的成本降低成本提供罕见的疾病基因检测,从而使这些关键测试更容易获得。链接学门户网站通过:•提供清晰,快速的遗传见解以更好地诊断•允许医生独立审查一组更广泛的遗传变异型•通过更好的专家Ramesh Hariharan博士,Ramesh Hariharan博士和Strand Life Signics的专家,我们的遗传诊断,我们的遗传学>“我们属于士兵”>“我们是属于遗传学的,我们都可以改善产前诊断,我们可以诊断出来。我们将继续创新并投资于诸如Strandomics门户网站之类的工具,以赋予医生与罕见疾病作斗争的能力。”关于Strand Life Sciences:Reliance Industries Limited的子公司Strand Life Sciences是一家基于基因组学的研究和诊断公司,将生物信息学中的长期记录与最先进的实验室分析和庞大的医院合作伙伴网络相结合,以推动新一代的患者护理Strand的客户包括全球医疗工具,诊断和制药公司。Strand还是印度基因组测试的先驱,其悠久的全球生物信息学记录使其成为肿瘤学,罕见疾病,妇女健康和传染病领域中基因组诊断最受信任的公司。此外,Strand正在积极扩展到消费者基因组学,使具有个性化见解的人能够对其基因组成。有关更多信息,请访问https://strandls.com

补贴建议单元报告零氢基金净4的链条
1的第2部分第1章要求公共权力在决定提供补贴之前考虑补贴的控制原则,能源和环境原则。公共当局不得授予补贴,除非这与这些原则一致。该法案第2部分的第2章禁止提供某些补贴,并且与某些其他类别的补贴有关,就可以遵守公共当局必须遵守的许多要求。2补贴咨询单元的补贴控制功能-Gov.uk(www.gov.uk)3其他链为其他类型的氢生产提供了支持:链1:DEVEX(开发支出),用于饲料研究和饲料后成本。Strand 2:不需要通过氢业务模型需要收入支持的项目的资本支出(资本支出)。strand 3:非CCUS启用项目的资本支出,这些项目也需要通过氢业务模型进行收入支持。

可以合成本身和互补链的聚合酶核酶
图1。三个小聚合酶核酶基序的发现和进化。(a)选择构造的格式用于初始选择回合(回合1至3或1至5),库是通过柔性链接器链接到杂交标签的六聚体标签的。生物素化引物可以捕获活性连接酶(在图中进行了详细描述S1-S2)。(b)在后期回合中使用的选择构建体的格式(3至11或5至11),需要三磷酸化的三核苷酸(Triplet)底物的聚合。在选择过程中,三胞胎(xxx)的序列(xxx)和由模板(x'x'x')编码的三重态数(y)在选择过程中变化(表S1中的详细信息)。(c)序列和预测从显示迭代三重三重连接的库中发现的三个核酶的二级结构,即三重酶聚合酶活性。在绿色中,源自随机库部分的核苷酸。在灰色的核苷酸中,源自恒定区域(接头和引物结合位点)。(d)在(b)中显示的(c)中显示的核酶的迭代三重聚会聚合,带有xxx = gcg和x'x'x'= cgc,y = 3。反应条件:50 nm核酶 - 基底,50 nm引物BCY3P10GA,50 nm模板T6FP10GAGCG3,5μMPPPGCG三胞胎,0.05%Tween 20,200 mm Kcl,50 mm Kcl,50 mm mgcl 2,50 mm mgcl 2,50 mm ches-koh,ph 9,3天,3天,以-77°°°°°°°°核酶与模板杂交。(E)序列和预测源自1-40克隆的QT51核酶的二级结构。黑色圆圈表示从1-40个祖先序列突变的6个残基;三角形表示2-核苷酸缺失。(f)60核苷酸序列的合成,该序列由CGU三重态的20个重复组成。Reaction conditions: 0.25 μM primer F10, 0.25 μM template tP10CGU20, 0.25 μM ribozyme, 10 μM pppCGU triplet, QT51 in 0.05% Tween 20, 50 mM MgCl 2 , 50 mM CHES-KOH, pH 9, 5TU+t1.5 in 200 mM MgCl 2 , 50 mM Tris-Cl, pH 8.3, 2 weeks在-7°C冷冻。核酶未与模板杂交。

双链到单链的转变在 DNA 折纸纳米结构中引起力和运动
除静态纳米结构外,DNA纳米技术还能构建动态和自主开关。[18] 这些动态开关的操作可分为两大类:第一,通过分子相互作用操作;第二,通过外部刺激操作。用于控制纳米尺度运动的主要分子相互作用是DNA杂交(主要是立足点介导的链置换)和碱基堆积。由分子相互作用控制的此类运动的例子包括可重构等离子体装置、[19] 铰链、[20,21] 镊子、[18,22] 旋转装置、[23–26] 助行器、[27] 药物载体 [28,29] 和对分子或纳米颗粒进行分选的机器人。[30,31] 作为驱动机制的其他分子相互作用包括靶分子结合 [32,33] 和适体 [28,29] 以及核小体相互作用。 [34] 通过任何分子相互作用进行的操作(包括上述所有机制)具有可控分子识别和特异性的优点。 然而,操作速度受到分子扩散和相互作用动力学的限制,因此通常非常慢。 值得注意的是,已经开发出多种方法来提高动态 DNA 装置的响应速度。 另一方面,外部刺激如光、[35,36] 温度、[37] 离子、[11,23] pH、[38–40] 和电场 [21,41] 通常能够使操作速度提高很多个数量级。[41] 例如,Karna 等人利用相邻纳米结构域之间可逆的、pH 依赖性的 i-基序形成来促进卷曲 DNA 纳米弹簧的驱动,进而通过整合素偶联影响培养细胞的运动性。 [40] 然而,我们在此称之为外部刺激的任何一种,都存在着整体作用的局限性,而且缺乏分子识别所能提供的特异性。
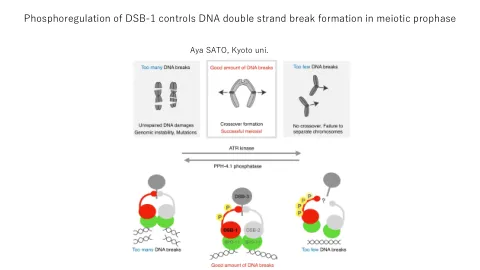

DNA双链断裂导致染色体染色单体
。cc-by-nc-nd 4.0国际许可证(未经同行评审证明)获得的是作者/资助者,他授予Biorxiv授予Biorxiv的许可,以永久显示预印本。它是制作

前导链和滞后链模板中的 DNA 缺口均可引发断裂诱导的复制
。CC-BY 4.0 国际许可下可用(未经同行评审认证)是作者/资助者,他已授予 bioRxiv 永久展示预印本的许可。它是此预印本的版权持有者此版本于 2023 年 12 月 22 日发布。;https://doi.org/10.1101/2023.12.22.573028 doi:bioRxiv 预印本