XiaoMi-AI文件搜索系统
World File Search SystemtRNA


ADAT:在 tRNA 编辑中的作用及其与疾病的相关性
摘要 转移 RNA (tRNA) 在蛋白质生物合成中起着核心作用。转录后 RNA 修饰影响 tRNA 的功能和稳定性。在这些修饰中,RNA 编辑是生命三个领域中广泛存在的 RNA 修饰。作用于 tRNA 的腺苷脱氨酶 (ADAT) 家族的蛋白质是在 20 多年前发现的。它们在 tRNA 成熟过程中催化腺苷脱氨为肌苷 (A - 到 - I) 或胞苷脱氨为尿苷 (C - 到 - U)。研究最多的例子是原核或真核 tRNA 反密码子中 tRNA 摆动位置的 TadA 或 ADAT2 / 3 介导的 A - 到 - I 转换。本综述提供了有关不同生命领域中 tRNA 的 A 到 I 和 C 到 U 编辑的详细信息,介绍了有关 DNA 编辑的 ADAT 的最新发现,最后评论了 ADAT3 基因突变与智力障碍之间的关联。

tRNA合成酶增强遗传代码扩展
使用正交翻译系统(OTSS)是通过在遗传密码中添加非经典氨基酸(NCAA)来产生非天然蛋白质的最有效方法。在寻求扩展底物特异性时,常规方法始于(超 - )稳定酶,能够承受由于必要突变而导致的结构变化。然而,我们在这里从发展以应对不稳定性的酶开始,从而占据根本不同的位置,因此可能对突变表现出更大的弹性。通过工程化甲烷菌Coides Burtonii的精神病(“冷”)OTS,我们开发了常用的中嗜和热嗜热系统的替代方法。即使在非常低的NCAA浓度下,这种OT在体内效率和滥交方面都显示出显着的特性。鉴于适用的寄主生物的广泛范围,我们预计冷酷将极大地促进扩展的遗传密码从学科转变为高价值化学驱动的生物技术。

tRNA 疗法在初创企业中崭露头角
lltrna 于 2021 年 11 月成立,自称是“世界上第一家 tRNA 平台公司”。通过设计转移 RNA 分子(蛋白质合成的细胞信使),这家初创公司获得了 5000 万美元的初始融资,旨在解决可能引发各种疾病的错误蛋白质生产机制。但 Alltrna 并不是唯一一家追求基于 tRNA 的疗法的公司。ReCode Therapeutics、Shape Therapeutics 和 Tevard Biosciences 都先于它而来;随着 hC Bioscience 的到来,该领域继续扩大,这家初创公司于 2 月底以 2400 万美元的融资从隐身模式中脱颖而出,计划用 tRNA 对抗癌症和罕见疾病。所有这些公司都至少在一定程度上专注于设计 tRNA 以绕过过早停止信号并改为整合所需的氨基酸。这种过早终止密码子的作用就像句子中间放错的句号,会混淆信使 RNA (mRNA) 中编码的信息,约占所有遗传疾病的 11%。因此,从理论上讲,仅一个“抑制” tRNA 就可能治愈数千种不同的罕见遗传疾病,每种疾病都是由相同类型的截断“无意义”突变引起的,这些突变会导致基因表达错误。非营利性囊性纤维化基金会的研究和药物发现战略顾问 William Skach 表示:“如果可以安全地做到这一点,那么它真的为一整类新疗法打开了大门。”Alltrna 的创始 CEO 兼董事 Lovisa Afzelius 补充道:“它释放了一种能力,可以满足那些原本被完全忽视的患者群体的未满足需求。”然而,尽管 tRNA 技术在临床前具有诸多前景,但目前尚不确定该平台是否会胜过小分子“读通”药物,如 Translarna (ataluren),这是一种在欧洲和巴西获批用于治疗无义突变介导的杜氏肌营养不良症的药物。此外,正在开发的基因编辑策略也可能与 tRNA 药物相媲美。“我们还不知道抑制 tRNA 在体内的功效,”阿拉巴马大学伯明翰分校的分子遗传学家 Kim Keeling 指出,他仍在继续寻找具有抑制 tRNA 的化合物

针对氨酰 tRNA 合成酶进行抗疟药物开发
疟原虫引起的感染给世界上最贫穷的社区带来了巨大的负担。我们迫切需要具有新作用机制的突破性药物。作为一种经历快速生长和分裂的生物体,疟原虫恶性疟原虫高度依赖蛋白质合成,而蛋白质合成又需要氨酰基-tRNA 合成酶 (aaRS) 为 tRNA 充电相应的氨基酸。蛋白质翻译是寄生虫生命周期所有阶段所必需的;因此,aaRS 抑制剂具有全生命周期抗疟活性的潜力。本综述重点介绍了使用表型筛选、靶标验证和结构引导药物设计来识别有效的疟原虫特异性 aaRS 抑制剂的努力。最近的研究表明,aaRS 是一类 AMP 模拟核苷磺酰胺的易感靶标,这些靶标通过一种新颖的反应劫持机制靶向酶。这一发现开辟了生成不同 aaRS 的定制抑制剂的可能性,从而提供了新的药物线索。

促进遗传代码解释的tRNA修饰模式
对核苷酸三元组到氨基酸的遗传密码的解释是生命的基础。 这种解释是通过细胞TRNA实现的,每个人都通过其互补反密码子(位置34-36)读取三胞胎密码子,同时将充电至其3'端的氨基酸传递。 然后将这种氨基酸掺入核糖体蛋白质合成期间的生长多肽链中。 解释的质量和多功能性不仅可以通过密码子与年代的配对来确保,而且还通过在每个tRNA的位置34和37处的转录后修饰来确保,分别对应于对应于抗构型抗源代码的第一个位置的旋转核苷酸,并相对于抗代支的3''侧。 如何通过匹配的反密码子读取每个密码子,以及需要哪些修改,因此不能单独使用密码子 - 抗议配对来预测。 在这里,我们提供了一个易于访问的修改模式,该模式集成到遗传代码表中。 我们将重点放在革兰氏阴性细菌大肠杆菌作为模型上,这是为数不多的生物之一,其整个tRNA修饰和修饰基因都被鉴定和映射。 这项工作提供了一个重要的参考工具,该工具将促进蛋白质合成研究,这是细胞寿命的核心。对核苷酸三元组到氨基酸的遗传密码的解释是生命的基础。这种解释是通过细胞TRNA实现的,每个人都通过其互补反密码子(位置34-36)读取三胞胎密码子,同时将充电至其3'端的氨基酸传递。然后将这种氨基酸掺入核糖体蛋白质合成期间的生长多肽链中。解释的质量和多功能性不仅可以通过密码子与年代的配对来确保,而且还通过在每个tRNA的位置34和37处的转录后修饰来确保,分别对应于对应于抗构型抗源代码的第一个位置的旋转核苷酸,并相对于抗代支的3''侧。如何通过匹配的反密码子读取每个密码子,以及需要哪些修改,因此不能单独使用密码子 - 抗议配对来预测。在这里,我们提供了一个易于访问的修改模式,该模式集成到遗传代码表中。我们将重点放在革兰氏阴性细菌大肠杆菌作为模型上,这是为数不多的生物之一,其整个tRNA修饰和修饰基因都被鉴定和映射。这项工作提供了一个重要的参考工具,该工具将促进蛋白质合成研究,这是细胞寿命的核心。
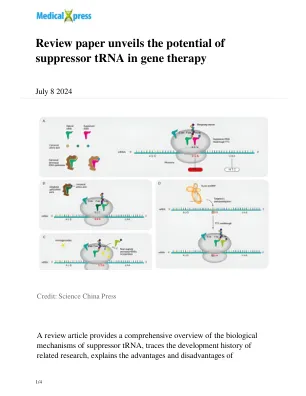
审查论文揭示了基因治疗中抑制器tRNA的潜力
最初,tRNA仅被认为是氨基酸的转运蛋白。通过发现抑制器tRNA发生了变化。1965年,Engelhardt等人。 实验表明,抑制器tRNA可以识别停止密码子和插入氨基酸,从而绕过翻译终止过程。 随后的研究进一步揭示了真核生物中抑制器tRNA的读取机理,并在基因治疗中显示了其潜力。1965年,Engelhardt等人。实验表明,抑制器tRNA可以识别停止密码子和插入氨基酸,从而绕过翻译终止过程。随后的研究进一步揭示了真核生物中抑制器tRNA的读取机理,并在基因治疗中显示了其潜力。

地球,创造生命的行星PCR机器或trna的简短历史
生命的起源和居住在包括我们自己的地球的物种的多样性是一个有趣的问题,它伴随着人类历史到当今。今天,数据库中基因组序列的令人惊叹的积累以及用于深入分析的无数生物信息学工具[1-3],清楚地表明,地球上生命的迷人生活史写在所有活物体的pangenome中。比较分歧DNA,RNA和/或蛋白质序列与共同祖先的比较,使我们能够将该历史的理论重建回到我们星球上的第一种自主细胞类型,该细胞在我们的星球上具有脂质包膜内具有核酸合成的核酸合成和核糖体的核酸群中的元素群体,该核酸属于蛋白质的概括[4] [5-7]。

使用氨基酰基-TRNA合成酶/tRNA对的遗传代码扩展的实用方法
摘要:遗传密码扩展(GCE)可以使非典型氨基酸(NCAA)的位点选择性掺入蛋白质中。GCE已大大提高,可用于在细胞内部创建生物策略手柄,监测和控制蛋白质,研究翻译后修饰和工程新蛋白质功能。自建立我们的实验室以来,我们的研究集中在使用氨基酰基-TRNA合成酶/tRNA(AARS/tRNA)对中GCE在蛋白质和酶工程中的应用。该主题已经进行了广泛的审查,毫无疑问,GCE是工程蛋白质和酶的强大工具。因此,对于这个年轻的教师问题,我们想对我们使用的方法以及我们在实验室中考虑的挑战进行更技术性的了解。自启动实验室以来,我们已经成功地使用了针对各种GCE应用量身定制的十二个新颖的AARS/tRNA对。但是,我们承认该领域即使对于专家也会构成挑战。因此,在此,我们提供了NCAA合并中的方法论,并提供了一些实践评论,并将重点放在挑战,新兴解决方案和令人兴奋的发展上。

初级保健中触发工具的开发和验证
氨基酰基-TRNA和GTP结合的翻译伸长因子EF-TU识别核糖体的A位点密码子取决于多肽(P)和出口(E)密码子位点中存在的密码子和TRNA物种。为了了解密码子环境如何影响tRNA结合的EF-TU识别密码子识别的效率,开发了一个遗传系统,可以通过慢速翻译密码子组合选择快速翻译。选择通过慢速翻译的UCA-UAC对,两侧是Histi Dine密码子,从而在必需的TRNA Leuz的D-STEM中分离了A25G碱基取代突变体,该突变体识别UUA和UUG亮氨酸密码子。Leuz(A25G)替换允许通过包括UCA密码子在内的所有密码子对进行更快的翻译。插入。这项工作是根据trpt tRNA中的Hirsh UGA非理性抑制剂G24a突变所做的,它提供了遗传证据,即通过伸长因子TU进行的GTP后水解校对校验拟合步骤可以通过TRNA物种铰链区域中的结构相互作用来控制。我们的结果支持一个模型,在该模型中,mRNA翻译中的tRNA弯曲成分允许EF TU时间增强其区分cognate和接近同名mRNA密码子之间的tRNA相互作用的能力。