XiaoMi-AI文件搜索系统
World File Search SystemtRNA

氨基酰基-TRNA合成酶的基因组重复导致
通过电解质选择作者揭示了分子量对糖化聚噻吩的混合传导的影响:Joshua Tropp,A,†Dilara Meli,B,B,†Ruiheng Wu,C Bohan Xu,B Samuel B.Hunt,D Jason D. Azoulay,D Bryan D. Paulsen,Jonathan Rivnay,A A A A A A A A A A A A A S NORTON WESTERN UNIXICANN,WESWESTERN UNIXICY,EVANSTON,伊利诺伊州伊利诺伊州60208,美国材料科学与工程系,伊利诺伊州伊利诺伊州伊利诺伊州60208,美国伊利诺伊州伊利诺伊州伊利诺伊州伊利诺伊州伊利诺伊州伊利诺伊州伊利诺伊州伊利诺伊州伊利诺伊州伊利诺伊州伊利诺伊州伊利诺伊州伊利诺伊州伊利诺伊州伊利诺伊州伊利诺伊州伊利诺伊州伊利诺伊州伊利诺伊州伊利诺伊州伊利诺伊州伊利诺伊州。州D州D。尚未彻底探索的一个重要特征是分子量对OMIEC性能的作用。在这项工作中,我们检查了一系列原型糖化的聚噻吩材料(P3meeet),系统地增加了有机电化学晶体管(OECTS)内的分子量 - 一种用于研究混合运输的普通测试型。我们发现,超出中间分子量的性能有所改善,但是,这种关系是电解质依赖性的。Operando分析表明,在NaCl中溶解在NaCl中的大量肿胀可能会因破坏结晶石电荷渗透而在NACL中造成巨大肿胀。这些发现证明了分子量和电解质组成的重要性,以增强OMIEC的性能。TOC ImageTOC Image通过在KTFSI中的操作揭示了分子量的作用,因为掺杂通过阳离子驱动而发生,从而防止了有害的肿胀并保持过敏性途径。

一种新型的噬菌体TRNA分解细菌的抗病毒防御...
一个联合研究小组,由治疗药物和疫苗开发中心的KIGA TSUNETARO组成(吉吉医学院医学院传染病学系的访问教授,访问教授,吉吉医学院医学院)等,已经巧妙地阐明了一种新的机制,可以巧妙地利用TRNA来促进细菌效应。这一发现可以鼓励进一步发展噬菌体疗法,并导致实现创新疗法的多药抗性细菌,这些疗法不太可能对传统的抗生素有效。

缬氨酸氨基酰基-TRNA合成酶促进黑色素瘤的耐药性
转移RNA动力学通过调节密码子特异性信使RNA翻译有助于癌症的发展。特定的氨基酰基-TRNA合成酶可以促进或抑制肿瘤发生。在这里我们表明valine氨基酰基-TRNA合成酶(VARS)是密码子偏置翻译重编程的关键参与者,该重编程是由于对靶向(MAPK)疗法在黑色素瘤中的抗性(MAPK)。患者衍生的MAPK治疗耐药性黑色素瘤中的蛋白质组会重新布线,偏向于valine的使用,并且与valine cognate trnas的上调以及VARS的表达和活性相吻合。引人注目的是,VAR敲低重新敏感了MAPK-耐药的患者衍生的黑色素瘤体外和体内。从机械上讲,VARS调节了富含Valine的转录本的使者RNA翻译,其中羟基酰基-COA脱氢酶mRNA编码用于脂肪酸氧化中的关键酶。耐药性黑色素瘤培养物依赖于脂肪酸氧化和羟基乙酰-COA脱氢酶在MAPK治疗后的生存。一起,我们的数据表明,VAR可能代表了治疗耐药性黑色素瘤的有吸引力的治疗靶点。


在酪酶基-TRNA合成酶基因YARS2相关的新生儿表型
人类遗传疾病通常是由复合杂合性突变引起的,其中突变基因的每个等位基因都具有不同的遗传病变。但是,由于缺乏适当的模型,对此类突变的研究受到阻碍。在这里,我们描述了在强制性酶二聚体中的复合异伴变体的动力学模型,该变体在一个单体中包含一个突变,而第二个单体中的另一个突变中包含一个突变。该酶由人YarS2编码用于Mito-trosyl-tRNA合成酶(MT-Tyrrs),该酶是氨基化酪氨酸到MT-TRNA Tyr的氨基酰基。yarS2是MT-氨基酰基-TRNA合成酶的基因的成员,其中致病性突变的疾病严重程度与酶活性之间的相关性有限。我们在YARS2中识别一对与新生儿死亡有关的化合物杂合变体。我们表明,虽然每个突变在MT-TYRR的同型二聚体中导致氨基酰化的最小缺陷,但反式跨性别的两个突变会协同降低酶活性,从而更大。因此,这种动力学模型准确地概括了疾病的严重程度,强调了其研究YARS2突变的效用及其对具有复合杂合突变的其他疾病的泛化潜力。

社论:转移RNA的合成生物学和治疗应用
在所有活细胞中,遗传代码定义了蛋白质编码基因中核酸序列与需要准确生产基因组中所有蛋白质所需的氨基酸序列之间的关系。氨基酰基-TRNA合成酶对转移RNA(TRNA)的氨基酰化是将氨基酸与TRNA守流量将氨基酸物理联系起来的关键步骤,从而决定了密码子与氨基酸的分配。由于它们在蛋白质合成中的核心作用,设计和合成的TRNA作为正交翻译系统的重要组成部分,旨在将非典型甚至不自然的氨基酸掺入细胞和无细胞系统中的蛋白质中。此外,最近的努力使用了正常的野生型或工程性TRNA来纠正引起人类疾病的遗传缺陷。由于11%的遗传遗传疾病是由过早的停止密码子引起的,因此废话抑制剂TRNA对tRNA疗法的应用越来越兴趣。我们认识到,TRNA的合成生物学和治疗应用都将依赖于胡说八道,在某些情况下,错过抑制tRNA,生成新型蛋白质或纠正遗传缺陷。因此,遗传学领域的研究主题具有遗传代码扩展和探索TRNA在合成生物学和医学应用中的应用中的作用的研究。

基于质谱的直接测序从头开始和多个RNA修饰的定量映射
摘要:尽管RNA的下一代测序(NGS)广泛使用,但多个RNA核苷酸修饰的同时直接测序和定量映射仍然具有挑战性。质谱(MS)的测序可以直接序列所有RNA修饰,而无需限于特定的测序,但是它需要很少有TRNA可以提供的完美MS梯子。在这里,我们描述了一种MS梯子互补测序方法(MLC-SEQ),该方法避免了完美的阶梯要求,从而可以在单核苷酸精度下对全长异质细胞TRNA进行全长异质细胞TRNA的测序。与基于NGS的方法(失去RNA修改信息)不同,MLC-Seq保留了RNA序列多样性和修改信息,揭示了新的详细的化学计量tRNA修饰谱及其在使用DealKylating酶ALKB治疗时进行的更改。也可以将其与参考序列结合使用,以提供对总TRNA样品中不同TRNA和修改的定量分析。MLC-Seq可以实现RNA修改的系统,定量和特定于位点的映射,从而揭示了TRNA的真正完整信息内容。■简介

通过体内超成名的氨基酰基-TRNA合成酶的定向演变
抽象的遗传密码扩展(GCE)已通过实现非经典氨基酸(NCAA)的位点掺入到蛋白质中,已成为生物学的关键工具。GCE的中心是正交氨基酰基-TRNA合成酶(AARS)/tRNA对的开发,其中工程的AARS识别所选的NCAA并将其充电到解码空白密码子的TRNA(例如,琥珀终止密码子)。许多正交的AARS/tRNA对涵盖了广泛的NCAA,这是通过定向进化产生的,但是标准策略通过标准策略的新AARS/TRNA对的演变仍然是一个劳动密集型的过程,通常会产生AARS/TRNA对,并产生副最好的NCAA NCAA INCAA Incorpiesies。在这项研究中,我们提出了一种发展AARS的策略,该策略利用Orthorep来推动其在酵母中的连续超女。我们在8个独立的AARS进化运动中展示了我们的战略,从4个不同的AARS/tRNA父母开始,针对7个不同的NCAA。我们观察到了多种新型AARS的快速演变,能够将13个NCAA的整体范围纳入响应于琥珀色密码子的蛋白质中。一些进化的系统达到了琥珀色密码子指定的NCAA依赖性翻译的效率,可与酵母中有义务密码子指定的天然氨基酸翻译相当。此外,我们发现了一个令人惊讶的AAR,它演变为自我调节自己的表达,以更大程度地依赖NCAA进行翻译。这些发现证明了由Orthorep驱动的AARS进化平台支持GCE技术持续增长的潜力。

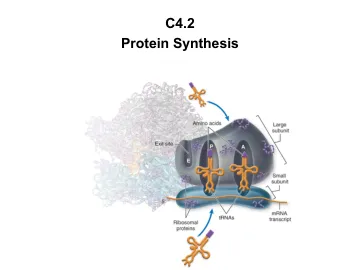
看到曙光——实时可视化蛋白质翻译
新的靶点空间 通过靶向 mRNA 来调节蛋白质表达已经是药物开发中一种经过验证的方法,如基于 RNAi 的疗法所示。然而,这些药物不是小分子,这严重限制了它们作为治疗策略的使用。用小分子靶向 mRNA 二级结构是一个新兴领域,但用小分子结合 mRNA 的治疗效果尚未得到验证。此外,这种方法无法实现组织选择性,因为此类药物会与目标 mRNA 在其表达的任何地方结合。Slonim 解释说,他的团队已经开发了一个使用荧光标记的转移 RNA (tRNA) 的高内涵筛选平台。tRNA 通过将特定氨基酸按照 mRNA 上的代码运输到将氨基酸链接到蛋白质的核糖体中发挥重要作用。“我们的想法是将荧光颜色附着在 tRNA 上,并将其转染到细胞中。当一对标记的 tRNA 最终位于核糖体上时,它会产生 FRET 信号,使我们能够实时看到蛋白质翻译的光,”他说。为了实现这一目标,该团队纯化并用荧光标签标记了人类所有 tRNA(46 种同工受体)(图 1)。他们设计了两个完整的 tRNA 库:一个标记有能量供体,另一个标记有能量受体(红色和绿色)。“这是产生 FRET 信号的绝佳机会。核糖体将两个 tRNA 放在一起,卸下氨基酸并构建蛋白质链。这是一个可重复的事件,发生在每个核糖体、每个细胞中——它可能是自然界中存在的最可重复的事件。我们可以在看起来像夜晚银河系的图像中看到这一现象,”他说。