XiaoMi-AI文件搜索系统
World File Search Systemtransduced

干细胞报告
精原干细胞 (SSC) 是生产转基因动物的资源。然而,对 SSC 的基因操作取得的成功有限。在这里,我们展示了通过慢病毒 (FV-LV) 使用融合蛋白 (F) 将基因有效转移到 SSC 中,融合蛋白是一种参与病毒体/细胞膜融合的仙台病毒 (SV) 包膜蛋白。FV-LV 比传统 LV 更有效地转导培养的 SSC。虽然感染 SV 的 SSC 无法产生后代,但用 FV-LV 转导的 SSC 具有生育能力。体内微注射表明 FV-LV 不仅可以穿透曲细精管的基底膜,还可以穿透血睾屏障,从而成功转导生精细胞和睾丸体细胞。用表达针对 Kit 或 Sycp3 的药物诱导型 CRISPR/Cas9 的 FV-LV 转染培养的 SSC 在移植和体内药物治疗后表现出精子发生受损。因此,FV-LV 为涉及 SSC 和精子发生的基因的功能分析提供了一种有效的方法。

CRISPR-Cas9 筛选用于目标识别。
图 2:PLX-4032 治疗后 sgRNA 计数分布的变化。log2 转换的 sgRNA 计数的箱线图和须线图(基线 - 抗生素选择后的转导细胞;DMSO 和 PLX-4032 - 筛选终点的细胞(治疗 14 天))。箱线图的范围从第一四分位数到第三四分位数,并以黑线显示中位数。此外,下部和上部相邻值显示为须线,异常值显示为圆圈。

DNA使用Omni Bead Ruptor 12珠磨机均质器从大麻sativa中提取DNA,以进行样品制备。
图4:a)MDA-MB-231细胞被绿色 - 肾上腺胶体易感病毒感染,并使用紫霉素选择了稳定的转导细胞。每周一次通过IVIS系统确定生物发光信号的强度六周。b)用红色葡萄糖和GFP标记的双重标记的MDA-MB-231细胞。可以通过两个FACS分析检测GFP报告基因表达。c)使用Nuance多光谱成像系统测量HT1080细胞中的GFP表达。

5.85.058 div>
背景Lyfgenia(Lovotibeglogene Autotemcel)是一种βA -T87Q-斑蛋白基因疗法,由含有疾病细胞疾病(SCD)的自体CD34 +细胞组成,其中含有造血性干细胞(HSC)(HSCS),该患者用BBBB305 LVV lvv sendy -bb305 lvv sendy -bb305 lvv sendy -bb305 lvv sendecode n lvv a a a a -a -t87q-globin。lyfgenia旨在一次性给药,以在患者自己的HSC中添加β-珠蛋白基因的修饰形式的功能副本。输注Lyfgenia后,转导的CD34+ HSC植入了骨髓中,并分化产生含有生物活性βA-A-T87Q-糖的红细胞,这些细胞将与α-蛋白结合以产生含有βA-T87Q-Globin(HBA-Globin(HBA T87Q))的功能性血红蛋白(HB)。HBA T87Q具有相似的氧结合亲和力和氧血红蛋白解离曲线至野生型HBA,可降低细胞内总血红蛋白S(HBS)水平,并旨在在空间上抑制HBS的聚合,从而限制了红细胞的镰刀(1)。调节状态FDA批准的指示:Lyfgenia是一种基于自体造血干细胞基因疗法,用于治疗12岁或以上患有镰状细胞疾病的患者和血管熟悉事件的病史(1)。使用的局限性:在与Lyfgenia进行治疗后,患有α-丘陵性特征(-α3.7/-α3.7)的患者可能患有贫血异常发育不良的贫血,可能需要慢性红细胞输血。lyfgenia尚未研究(1)。
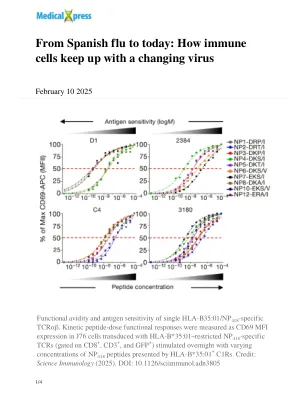
从西班牙流感到今天:免疫细胞如何跟上病毒的变化
单个 HLA-B35:01/NP 418 特异性 TCRαβ 的功能亲和力和抗原敏感性。动力学肽剂量功能反应通过 J76 细胞中的 CD69 MFI 表达来测量,这些细胞被 HLA-B*35:01 限制的 NP 418 特异性 TCR(门控 CD8 + 、CD3 + 和 GFP + )转导,并用不同浓度的 HLA-B*35:01 + C1R 呈递的 NP 418 肽刺激过夜。来源:Science Immunology (2025)。DOI:10.1126/sciimmunol.adn3805

服务表免疫肿瘤学:癌症免疫疗法革命刚刚开始
HBV小鼠模型:通过AAV载体转导HBV的免疫胜任动物,随着时间的推移,通过针对特定应用(直接抗病毒药或宿主靶向剂)定制的治疗方案,导致持续的病毒产物。疾病相关的终点包括:通过RT-PCR确定循环和肝脏病毒DNA和RNA;通过DDPCR和Southern印迹进行CCCDNA定量;分析病毒抗原HBEAG/ HBSAG;免疫浸润和细胞因子释放的分析;肝组织和AST/ALT评估的免疫历史学。

2022; 18(1): 426-440。doi: 10.7150/ijbs.64640 研究论文 抗多种自然杀伤亚型的人类间充质干细胞工程
具有治疗前景的间充质干细胞 (MSC) 通常会被宿主的先天免疫细胞(包括自然杀伤 (NK) 细胞)迅速清除。人们已努力生成免疫逃逸的人类胚胎干细胞 (hESC),其中通过缺陷的 β -2-微球蛋白 (B2M)(人类白细胞抗原 (HLA) I 类的共同单位)逃避 T 细胞免疫,并通过 HLA-E 或 -G 的异位表达抑制 NK 细胞。然而,NK 亚型在接受者之间甚至在不同的病理状态下都不同。有必要剖析和优化免疫逃逸细胞对 NK 亚型的功效。在这里,我们首先生成 B2M 敲除 hESC 并将其分化为 MSC(EMSC),并发现仅当通过可诱导慢病毒系统以剂量依赖性方式转导时,表达 HLA-E 和 -G 的 B2M -/- EMSC 才会出现 NK 抗性,而当它们插入安全港时则不会出现。在转导的 EMSC 中同时高水平表达的 HLA-E 和 -G 抑制了三种主要 NK 亚型,包括 NKG2A + / LILRB1 + 、NKG2A + / LILRB1 - 和 NKG2A - / LILRB1 + ,并且 IFN- γ 启动进一步增强了这种抑制作用。因此,这项研究设计了对多种 NK 亚型具有抗性的 MSC,并强调当转基因用于赋予宿主细胞新效应时,剂量很重要,尤其是对于治疗细胞逃避免疫排斥而言。

长期在小鼠成纤维细胞植入物中逆转录病毒介导的基因转移的体内表达
当前的基因治疗模型涉及逆转录病毒介导的遗传材料转移到源自各种体细胞组织的细胞中,包括造血系统的细胞,成纤维细胞,肝细胞,内皮细胞和成肌细胞(1、2)。我们先前已经描述了一种通过小鼠皮肤成纤维细胞逆转录病毒感染的基因产物传递方法(3)。我们先前在成纤维细胞研究中使用的转导基因是人和狗因子IX cDNA(3,4)。尽管在组织培养中可以实现高水平的持续性,而当在啮齿动物的同种异体移植中移植时,这些成纤维细胞仅在短时间内就产生了大量因子IX(3,5)。从理论上讲,体内表达的短期可能归因于不同的因素:(i)宿主对外源性因子IX的免疫反应; (ii)移植后外国细胞的破坏; (IIM)一旦将转导细胞移植到动物的转移基因的转录基因转录的特异性下降。已经表明(3,5),植入改良的成纤维细胞后,对人类因子IX的抗体存在,这至少可以解释,部分原因是第IX因子的短期。在这项工作中,使用不同的启动子来控制8-半乳糖苷酶的表达,我们证明,在组织培养中,长期表达可以轻松获得,但指导感兴趣基因转录的启动子的类型可能是决定体内长期表达的关键因素之一。

de从设计的PMHC粘合剂促进T细胞诱导癌细胞杀死
通过T细胞受体(TCRS)对CD8 + T细胞对细胞内抗原的识别对于适应性免疫是至关重要的,可以针对感染和癌症产生反应。最近批准TCR基因编辑的T细胞用于癌症治疗,证明了使用PMHC识别消除癌症的治疗优势。但是,从患者材料中识别和选择TCR是复杂的,并且受使用的捐赠者的TCR库的影响。为了克服这些局限性,我们在这里提出了一个快速且坚固的DE NOVEN-DE DE平台,该平台利用了最新的生成模型,包括RfDiffusion,Proteinmpnn和Alphafold2,以靶向癌症相关PMHC Complex,NY-ESO-1(NY-ESO-1(157-165) /HLA-A-HALA-A*02.02.02.02通过将其纳入硅交叉铺设和分子动力学模拟中,我们增强了特异性筛选,以最大程度地减少脱靶相互作用。我们确定了一种MIBD,该MIBD对NY-ESO-1-衍生的肽Sllmwitqc具有很高的特异性,其中HLA-A*02:01和哺乳动物显示分析中的最小交叉反应性。我们通过将其整合到嵌合抗原受体中,进一步证明了该MIBD的治疗潜力,作为免疫介导的杀伤剂(Bikes)的从头粘合剂(自行车)。bike-与非转导的对照相比,有效地有效地杀死了NY-ESO-1 +黑色素瘤细胞的T细胞,证明了这种方法在精确癌症免疫疗法中的希望。我们的发现强调了生成蛋白设计在加速高特异性PMHC靶向疗法方面的变革潜力。除了使用CAR-T应用程序,我们的工作流程为开发MIBD作为多功能工具而建立了基础,预示了精确免疫疗法的新时代。
